2.3. Обмен холестерола, поступление, синтез, выведение. Эфиры холестерола
II. Переваривание и всасывание пищевых липидов
С пищей в организм ежедневно поступает от 80 до 150 г липидов. Основную массу составляют жиры, наряду с глюкозой служащие главными источниками энергии. Хотя калорийность жиров значительно выше, чем углеводов (9 по сравнению с 4,7 ккал/моль), при рациональном питании жиры обеспечивают не более 30% от общего количества калорий, поступающих с пищей. Жидкие жиры (масла) содержат в своём составе полиеновые жирные кислоты, которые не синтезируются в организме; поэтому жидкие жиры должны составлять не менее одной трети жиров пищи. С липидами в организм поступают
379

Рис. 8-9. Холестерол и его эфиры.
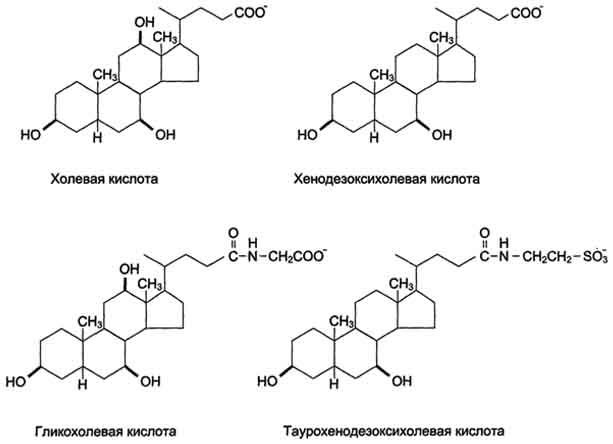
Рис. 8-10. Жёлчные кислоты.
380
и жирорастворимые витамины A, D, Е, К. Переваривание липидов пищи происходит в кишечнике. Основные продукты гидролиза (жирные кислоты и 2-моноацилглицеролы) после всасывания подвергаются ресинтезу и последующей упаковке в хиломикроны (ХМ) в клетках слизистой оболочки кишечника.
А. Эмульгирование жиров
Жиры составляют до 90% липидов, поступающих с пищей. Переваривание жиров происходит в тонком кишечнике, однако уже в желудке небольшая часть жиров гидролизуется под действием "липазы языка". Этот фермент синтезируется железами на дорсальной поверхности языка и относительно устойчив при кислых значениях рН желудочного сока. Поэтому он действует в течение 1-2 ч на жиры пищи в желудке. Однако вклад этой липазы в переваривание жиров у взрослых людей незначителен. Основной процесс переваривания происходит в тонкой кишке.
Так как жиры - нерастворимые в воде соединения, то они могут подвергаться действию ферментов, растворённых в воде только на границе раздела фаз вода/жир. Поэтому действию панкреатической липазы, гидролизующей жиры, предшествует эмульгирование жиров. Эмульгирование (смешивание жира с водой) происходит в тонком кишечнике под действием солей жёлчных кислот (рис. 8-11). Жёлчные кислоты синтезируются в печени из холестерола и сек-ретируются в жёлчный пузырь. Содержимое жёлчного пузыря - жёлчь. Это вязкая жёлто-зелёная жидкость, содержащая главным образом жёлчные кислоты; в небольшом количестве имеются фосфолипиды и холестерол. Жёлчные кислоты представляют собой в основном конъюгированные жёлчные кислоты: таурохолевую, гликохолевую и другие (см. выше рис. 8-10). После приёма жирной пищи жёлчный пузырь сокращается и жёлчь изливается в просвет двенадцатиперстной кишки. Жёлчные кислоты действуют как детергенты, располагаясь на поверхности капель жира и снижая поверхностное натяжение. В результате крупные капли жира распадаются на множество мелких, т.е. происходит эмульгирование жира. Эмульгирование приводит к увеличению площади поверхности раздела фаз жир/вода, что ускоряет гидролиз жира панкреатической липазой. Эмульгированию способствует и перистальтика кишечника.
Б. Гормоны, активирующие переваривание жиров
При поступлении пищи в желудок, а затем в кишечник клетки слизистой оболочки тонкого кишечника начинают секретировать в кровь пептидный гормон холецистокинин (панкреозимин). Этот гормон действует на жёлчный пузырь, стимулируя его сокращение, и на экзокринные клетки поджелудочной железы, стимулируя секрецию пищеварительных ферментов, в том числе панкреатической липазы. Другие клетки слизистой оболочки тонкого кишечника в ответ на поступление из желудка кислого содержимого выделяют гормон секретин. Секретин - гормон пептидной природы, стимулирующий секрецию бикарбоната (НСО3-) в сок поджелудочной железы.
В. Переваривание жиров панкреатической липазой
Переваривание жиров - гидролиз жиров панкреатической липазой. Оптимальное значение рН для панкреатической липазы ≈8 достигается путём нейтрализации кислого содержимого, поступающего из желудка, бикарбонатом, выделяющимся в составе сока поджелудочной железы:
Н+ + НСО3- → Н2СО3 → Н2О + СО2 ↑.
Выделяющийся углекислый газ способствует дополнительному перемешиванию содержимого тонкой кишки.
Панкреатическая липаза выделяется в полость тонкой кишки из поджелудочной железы вместе с белком колипазой. Колипаза попадает в полость кишечника в неактивном виде и частичным протеолизом под действием трипсина превращается в активную форму. Колипаза своим гидрофобным доменом связывается с поверхностью мицеллы эмульгированного жира. Другая часть молекулы способствует формированию такой конформации панкреатической липазы, при которой активный центр фермента максимально приближен к своим субстратам - молекулам жиров (рис. 8-12), поэтому скорость реакции гидролиза жира резко возрастает.
381
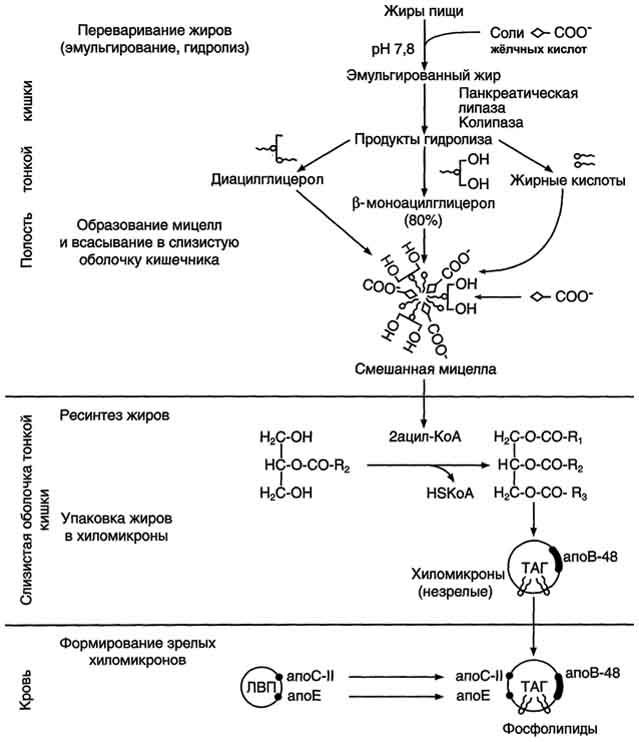
Рис. 8-11. Этапы поступления экзогенных жиров в организм.
382
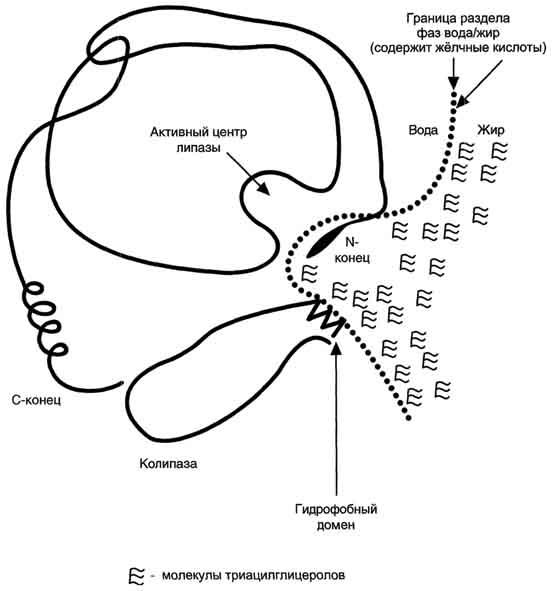
Рис. 8-12. Расположение панкреатической липазы и колипазы на границе раздела фаз вода/жир.
Панкреатическая липаза гидролизует жиры преимущественно в положениях 1 и 3 (рис. 8-13), поэтому основными продуктами гидролиза являются свободные жирные кислоты и 2-моноацилглицеролы (β-моноацилглицеролы).
Молекулы 2-моноацилглицеролов также обладают детергентными свойствами и способствуют эмульгированию жира.
Г. Переваривание других липидов
Кроме жиров, с пищей поступают фосфолипиды, эфиры холестерола, однако количество этих липйдов в составе пищи значительно меньше, чем жиров (≈10%).
Переваривание глицерофосфолипидов
В переваривании глицерофосфолипидов участвуют несколько ферментов, синтезирующихся в поджелудочной железе. Фосфолипаза А2 гидролизует сложноэфирную связь у второго атома углерода глицерола, превращая глицерофосфолипиды в соответствующие лизофосфолипиды. На рисунке 8-14 представлен пример гидролиза фосфатидилхолинов при переваривании.
383
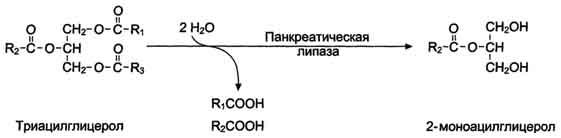
Рис. 8-13. Гидролиз триацилглицеролов панкреатической липазой.
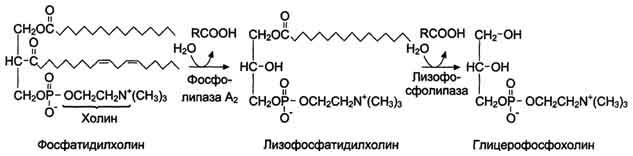
Рис. 8-14. Переваривание фосфатидилхолинов.
Фосфолипаза A2 секретируется в кишечник в виде профермента и активируется уже в полости кишечника путём частичного протеолиза. Для проявления активности фосфолипазы A2 необходимы ионы кальция.
Жирная кислота в положении 1 отщепляется под действием лизофосфолипазы, а глицерофосфохолин гидролизуется далее до глицерола, холина и фосфорной кислоты, которые всасываются. Лизофосфолипиды - эффективные эмульгаторы жира, ускоряющие его переваривание.
Переваривание эфиров холестерола
В составе пищи холестерол находится в основном в виде эфиров. Гидролиз эфиров холестерола происходит под действием холестеролэстеразы - фермента, который также синтезируется в поджелудочной железе и секретируется в кишечник (рис. 8-15). Продукты гидролиза (холестерол и жирные кислоты) всасываются в составе смешанных мицелл.
Д. Переваривание жира у грудных детей
У грудных детей и детей младшего возраста основной пищей служит молоко. Молоко содержит жиры, в состав которых входят в основном жирные кислоты с короткой и средней длиной алифатических цепей (4-12 атомов углерода). Жиры в составе молока находятся уже в эмульгированном, смешанном с водой виде, поэтому они сразу же доступны для гидролиза ферментами. На жиры молока в желудке детей действует липаза, которая синтезируется в железах языка (липаза языка). Кроме того, в желудке детей грудного и младшего возраста вырабатывается желудочная липаза, которая активна при нейтральном значении рН, характерном для желудочного сока детей, и не активна у взрослых (рН желудочного сока -∼1,5). Эта липаза гидролизует жиры, отщепляя, в основном, жирные кислоты у третьего атома углерода глицерола. Далее гидролиз жиров молока продолжается в кишечнике под действием панкреатической липазы. Жирные кислоты с короткой цепью, как водорастворимые, всасываются частично уже в желудке. Остальные жирные кислоты всасываются в тонком кишечнике. Для детей грудного возраста основным источником энергии являются жиры, в то время как у взрослых людей при нормальном питании основным источником энергии служит глюкоза.
384
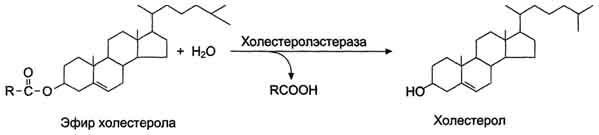
Рис. 8-15. Гидролиз эфиров холестерола в тонкой кишке.
Вследствие этого нарушение переваривания и всасывания жиров у детей более опасно, чем у взрослых.
Е. Всасывание продуктов гидролиза липидов в тонком кишечнике. ресинтез жиров
Образование смешанных мицелл и всасывание продуктов гидролиза
Продукты гидролиза липидов - жирные кислоты с длинным углеводородным радикалом, 2-моноацилглицеролы, холестерол, а также соли жёлчных кислот образуют в просвете кишечника структуры, называемые смешанными мицеллами. Смешанные мицеллы построены таким образом, что гидрофобные части молекул обращены внутрь мицеллы, а гидрофильные - наружу, поэтому мицеллы хорошо растворяются в водной фазе содержимого тонкой кишки. Стабильность мицелл обеспечивается в основном солями жёлчных кислот. Мицеллы сближаются со щёточной каймой клеток слизистой оболочки тонкого кишечника, и липидные компоненты мицелл диффундируют через мембраны внутрь клеток. Вместе с продуктами гидролиза липидов всасываются жирорастворимые витамины A, D, Е, К и соли жёлчных кислот. Наиболее активно соли жёлчных кислот всасываются в подвздошной кишке. Жёлчные кислоты далее попадают через воротную вену в печень, из печени вновь секретируются в жёлчный пузырь и далее опять участвуют в эмульгировании жиров. Этот путь жёлчных кислот называют "энтерогепатическая циркуляция". Каждая молекула жёлчных кислот за сутки проходит 5- 8 циклов, и около 5% жёлчных кислот выделяется с фекалиями.
Всасывание жирных кислот со средней длиной цепи, образующихся, например, при переваривании липидов молока, происходит без участия смешанных мицелл. Эти жирные кислоты из клеток слизистой оболочки тонкого кишечника попадают в кровь, связываются с белком альбумином и транспортируются в печень.
Ресинтез жиров в слизистой оболочке тонкого кишечника
После всасывания продуктов гидролиза жиров жирные кислоты и 2-моноацилглицеролы в клетках слизистой оболочки тонкого кишечника включаются в процесс ресинтеза с образованием триацилглицеролов (рис. 8-16). Жирные кислоты вступают в реакцию этерификации только в активной форме в виде производных коэнзима А, поэтому первая стадия ресинтеза жиров - реакция активации жирной кислоты:
HS КоА + RCOOH + АТФ → R-CO ~ КоА + АМФ + Н4Р2О7.
Реакция катализируется ферментом ацил-КоА-синтетазой (тиокиназой). Затем ацил~КоА участвует в реакции этерификации 2-моноацилглицерола с образованием сначала диацилгли-церола, а затем триацилглицерола. Реакции ресинтеза жиров катализируют ацилтранеферазы.
В реакциях ресинтеза жиров участвуют, как правило, только жирные кислоты с длинной углеводородной цепью. В ресинтезе жиров участвуют не только жирные кислоты, всосавшиеся из кишечника, но и жирные кислоты, синтезированные в организме, поэтому по составу ре-синтезированные жиры отличаются от жиров, полученных с пищей. Однако возможности "адаптировать" в процессе ресинтеза состав пищевых жиров к составу жиров организма человека ограничены, поэтому при поступлении с пищей жиров с необычными жирными кислотами,
385
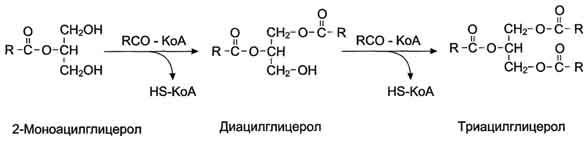
Рис. 8-16. Ресинтез жиров в клетках слизистой оболочки тонкой кишки.
например бараньего жира, в адипоцитах появляются жиры, содержащие кислоты, характерные для бараньего жира (насыщенные разветвлённые жирные кислоты). В клетках слизистой оболочки кишечника происходит активный синтез глицерофосфолипидов, необходимых для формирования структуры липопротеинов - транспортных форм липидов в крови.
Образование эфиров холестерола
В клетках слизистой оболочки тонкой кишки всосавшиеся молекулы холестерола также превращаются в эфиры путём взаимодействия с ацил-КоА (рис. 8-17). Эту реакцию катализирует ацилхолестеролацилтрансфераза (АХАТ). От активности этого фермента зависит скорость поступления экзогенного холестерола в организм.
В клетках эпителия тонкой кишки из жиров, образовавшихся в результате ресинтеза, а также из эфиров холестерола, жирорастворимых витаминов, поступивших с пищей, формируются ли-попротеиновые комплексы - хиломикроны (ХМ). ХМ далее доставляют жиры в периферические ткани.
Нарушения переваривания и всасывания жиров. Стеаторея
Нарушение переваривания жиров может быть следствием нескольких причин. Одна из них - нарушение секреции жёлчи из жёлчного пузыря при механическом препятствии оттоку жёлчи. Это состояние может быть результатом сужения просвета жёлчного протока камнями, образующимися в жёлчном пузыре, или сдавлением жёлчного протока опухолью, развивающейся в окружающих тканях. Уменьшение секреции жёлчи приводит к нарушению эмульгирования пищевых жиров и, следовательно, к снижению способности панкреатической липазы гидролизовать жиры.
Нарушение секреции сока поджелудочной железы и, следовательно, недостаточная секреция панкреатической липазы также приводят к снижению скорости гидролиза жиров. В обоих случаях нарушение переваривания и всасывания жиров приводит к увеличению количества жиров в фекалиях - возникает стеа-торея (жирный стул). В норме содержание жиров в фекалиях составляет не более 5%. При стеаторее нарушается всасывание жирорастворимых витаминов (A, D, E, К) и незаменимых жирных кислот, поэтому при длительно текущей стеаторее развивается недостаточность этих незаменимых факторов питания с соответствующими клиническими симптомами (см. раздел 3). При нарушении переваривания жиров плохо перевариваются и вещества нелипидной природы, так как жир обволакивает частицы пищи и препятствует действию на них ферментов.
studfiles.net
- транспорт холестерола - Биохимия
Транспорт холестерола и его эфиров осуществляется липопротеинами низкой и высокой плотности.
Липопротеины высокой плотности
Общая характеристика
- образуются в печени de novo, в плазме крови при распаде хиломикронов, некоторое количество в стенке кишечника,
- в составе частицы примерно половину занимают белки, еще четверть фосфолипиды, остальное холестерин и ТАГ (50% белка, 7% ТАГ, 13% эфиров ХС, 5% свободного ХС, 25% ФЛ),
- основным апобелком является апо А1, содержат апоЕ и апоСII.
Функция
- Транспорт свободного ХС от тканей к печени.
- Фосфолипиды ЛПВП являются источником полиеновых кислот для синтеза клеточных фосфолипидов и эйкозаноидов.
Метаболизм
1. Синтезированный в печени ЛПВП (насцентный или первичный) содержит в основном фосфолипиды и апобелки. Остальные липидные компоненты накапливаются в нем по мере метаболизма в плазме крови.
2-3. В плазме крови насцентный ЛПВП переходит в ЛПВП3 (условно его можно назвать "зрелый") и тесно взаимодействует с ЛПНП и ЛПОНП. Получая от них свободный холестерин, ЛПВП3 в обмен отдают эфиры ХС, образованные благодаря переносу жирной кислоты от фосфатидилхолина на холестерин.
Внутри ЛПВП активно протекает реакция при участии лецитин:холестерол-ацилтрансферазы (ЛХАТ-реакция). В этой реакции остаток полиненасыщенной жирной кислоты переносится от ФХ на свободный ХС с образованием лизофосфатидилхолина (лФХ) и эфиров ХС. Лизофосфатидилхолин остается внутри ЛПВП, эфир холестерина отправляется в ЛПНП.
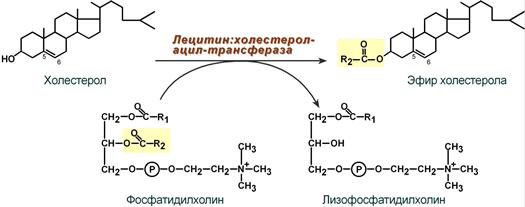 Реакция этерификации холестерола при участии лецитин:холестерол-ацилтрансферазы
Реакция этерификации холестерола при участии лецитин:холестерол-ацилтрансферазы 4. Взаимодействуя с ЛПОНП и ХМ, ЛПВП получают МАГ и ДАГ и отдают им апоЕ- и апоСII-белки.
5. При непосредственном контакте с клеточными мембранами или при участии специфических транспортных белков получают от них свободный ХС.
6. Взаимодействует с мембранами клеток, отдает часть фосфолипидной оболочки, доставляя таким образом полиеновые жирные кислоты в клетки.
7. Накопление свободного ХС, МАГ и ДАГ, лизоФХ и утрата фосфолипидной оболочки преобразует ЛПВП3 в ЛПВП2 (остаточный, ремнантный). Последний захватывается гепатоцитами при помощи апоА-1-рецептора и разрушается.
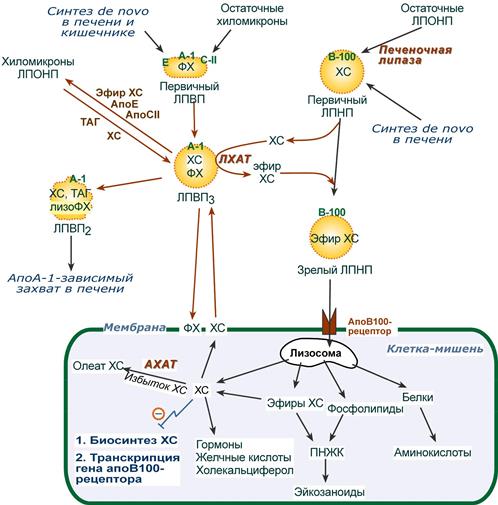 Транспорт холестерола и его эфиров в организме
Транспорт холестерола и его эфиров в организме Липопротеины низкой плотности
Общая характеристика
- образуются в гепатоцитах de novo и в сосудистой системе печени под воздействием печеночной ТАГ-липазы из ЛПОНП,
- в составе преобладают холестерол и его эфиры, около половины занимают белки и фосфолипиды (25% белки, 7% триацилглицеролы, 38% эфиров ХС, 8% свободного ХС, 22% фосфолипидов),
- основным апобелком является апоВ-100,
- нормальное содержание в крови 3,2-4,5 г/л,
- самые атерогенные.
Функция
1. Транспорт холестерола в клетки, использующие его для реакций синтеза половых гормонов (половые железы), глюкокортикоидов и минералокортикоидов (кора надпочечников), холекальциферола (кожа), утилизирующие ХС в виде желчных кислот (печень).
2. Транспорт полиеновых жирных кислот в виде эфиров ХС в некоторые клетки рыхлой соединительной ткани (фибробласты, тромбоциты, эндотелий, гладкомышечные клетки), в эпителий гломерулярной мембраны почек, в клетки костного мозга, в клетки роговицы глаз, в нейроциты, в базофилы аденогипофиза.
Особенностью всех этих клеток является наличие лизосомальных кислых гидролаз, расщепляющих эфиры ХС. У других клеток таких ферментов нет.
Клетки рыхлой соединительной ткани активно синтезируют эйкозаноиды. Поэтому им необходим постоянный приток полиненасыщенных жирных кислот (ПНЖК), что осуществляется либо переходом фосфолипидов от оболочки ЛПВП в мембраны клеток, либо поглощением ЛПНП, которые несут ПНЖК в составе эфиров холестерола.
Обмен
1. В крови первичные ЛПНП взаимодействуют с ЛПВП, отдавая свободный ХС и получая этерифицированный. В результате в них происходит накопление эфиров ХС, увеличение гидрофобного ядра и "выталкивание" белка апоВ-100 на поверхность частицы. Таким образом, первичный ЛПНП переходит в зрелый.
2. На всех клетках, использующих ЛПНП, имеется высокоафинный рецептор, специфичный к ЛПНП – апоВ-100-рецептор. При взаимодействии ЛПНП с рецептором происходит эндоцитоз липопротеина и его лизосомальный распад на составные части – фосфолипиды, белки (и далее до аминокислот), глицерол, жирные кислоты, холестерол и его эфиры.
- ХС превращается в гормоны или включается в состав мембран,
- излишки мембранного ХС удаляются с помощью ЛПВП,
- принесенные с эфирами ХС ПНЖК используются для синтеза эйкозаноидов или фосфолипидов.
- при невозможности удалить ХС часть его этерифицируется с олеиновой кислотой ферментом ацил-SКоА:холестерол-ацилтрансферазой (АХАТ),
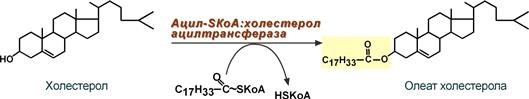 Синтез олеата холестерола при участии ацил-SKoA-холестерол-ацил-трансферазы
Синтез олеата холестерола при участии ацил-SKoA-холестерол-ацил-трансферазы 3. Около 50% ЛПНП взаимодействуют с апоВ-100-рецепторами гепатоцитов и примерно столько же поглощаются клетками других тканей.
Вы можете спросить или оставить свое мнение.
biokhimija.ru
2.3. Обмен холестерола, поступление, синтез, выведение.
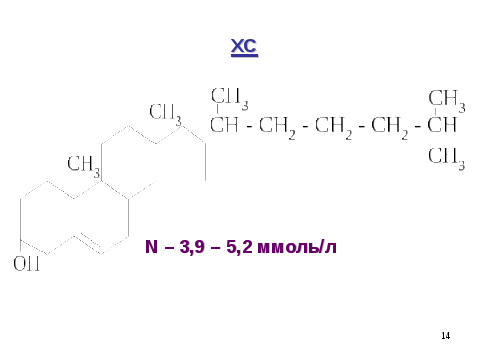
Холестерол – стероид, характерный только для животных организмов. 50% синтезируется в печени, 15-20% в тонком кишечнике, остальной – в коже, коре надпочечников, половых железах. В сутки синтезируется 1г холестерола. С пищей поступает 300-500мг.
N в крови = 3,9-5,2 ммоль/л
У новорожденных ХС – 1,3-2,6 ммоль/л
В 12-14 лет – достигает нормы.
Биологическая роль.
Входит в состав клеточных мембран, влияя на их свойства.
Субстрат для синтеза желчных кислот, стероидных гормонов, витамин Д3.
Поддерживает тургор кожи.
Реакция синтеза ХС происходит в цитолизе клеток, это один из самых длинных метаболических путей в организме человека, включает около 100 последовательных реакций.

2.4. Пути превращения холестерола в клетке: окисление в желчные кислоты, эстерификация, дегидрирование. Регуляция метаболизма холестерола.
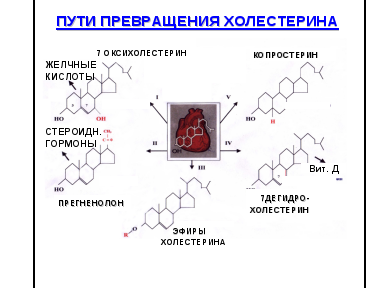
Основной путь превращения ХС – это его окисление. 80% от всего количества окисляется в печени в желчные кислоты, 3% в стероидные гормоны. При реакциях окисления в молекуле ХС появляются полярные группы, гидроксилы, карбоксилы, повышается его растворимость в воде, что способствует выведению из организма.
Второй путь превращения ХС в организме – это образование эфиров ХС (эстерификация). Эфиры ХС составляют около 10% от общего количества ХС в организме. При этом растворимость уменьшается и это приводит к накоплению в организме. У новорожденных относительно низкий коэффициент эстерификации по сравнению с более старшим возрастом детей = 0,58-1 ммоль/л, что связано с дефицитом полиненасыщенных жирных кислот.
Дегидрирование. В положении 7, 8 образуются двойные связи, что приводит к образованию 7-дегидрохолестерола (предшественника витамина Д3)
Выведение холестерола.
Чтобы поддерживать постоянный уровень его в организме необходимо выводить 1,5г в сутки. 1г в сутки окисляется в желчные кислоты, 200-300мг в сутки с калом в виде копростанола, 100мг со слущенным эпителием, 40 мг идет на синтез стероидных гормонов, с мочой 1-2мг в сутки.
Регуляция метаболизма холестерола.
Ключевую роль в регуляции метаболизма холестерина играют рецепторы ЛПНП. Впервые эти рецепторы были обнаружены на клетках кожи человека учеными Браун и Гольштейн в 1973г в Далласе (Техасском медицинском центре). За это открытие они получили нобелевскую премию по медицине в 1985г.
ЛПНП поставляют холестерин в различные клетки. Рецепторы, связывая белково-липидные комплексы ЛПНП не только обеспечивают клетки холестерином, но и снижают уровень ЛПНП в крови. Ясно, чем больше рецепторов ЛПН на поверхности клетки и чем эффективнее они взаимодействуют с ЛПН, тем быстрее холестерин удаляется из крови.
Происходит это в следующие этапы:
Рецепторы (по химической природе – гликопротеины) на плазматической мембране клеток узнают те ЛП, в состав которых входит апопротеин В-100 и связывается с ними.
Комплекс рецептор-ЛПНП проникает в клетку путем эндоцитоза, образуя пузырек-эндосому.
Эндосомы сливаются с лизосомами. Под действием ферментов лизосом белковая часть расщепляется до свободных аминокислот. Эфиры холестерина расщепляются кислой липазой до св. ХС.
Накапливающийся в клетках ХС оказывает тройное действие на метаболизм.
1. Тормозит синтез ХС самой клеткой, ингибируя по механизму обратной связи фермент ОмГ-редуктазу.
2. Индуцирует процесс образования эфиров ХС в цитоплазме клетки (активирует ЛХАТ) лецитинацетилхолестеролтрансфераза.
3. Избыток ХС по принципу обратной связи подавляет образование новых рецепторов ЛПНП. В результате количество рецепторов уменьшается и клетка начинает связывать меньше ЛПНП.
В регуляции синтеза рецепторов ЛПНП участвуют: инсулин, Т3, половые гормоны, которые увеличивают их количество, а глюкагон подавляет.
Эффекты инсулина и Т3 могут объяснить механизм гиперхолестеринемии, риск атеросклероза при сахарном диабете или при гипотиреозе.
studfiles.net
Общий холестерин и его фракции
Холестерин — одноатомный спирт, в молекуле которого имеется ядро циклопентанпергидрофенатрена. Он является компонентом клеточных мембран, предшественником при синтезе желчных кислот, стероидных гормонов (глюкокортикоидов, альдостерона, половых гормонов), витамина D, обнаруживается во всех тканях и жидкостях организма как в свободном состоянии, так и в виде эфиров с жирными кислотами, преимущественно с линолевой (около 10% всего холестерина). Синтез холестерина происходит во всех клетках организма. Основными транспортными формами в крови являются α‑, β‑ и преβ‑липопротеины (или, соответственно, липопротеины высокой, низкой и очень низкой плотности). В плазме крови холестерин находится главным образом в форме сложных эфиров (60‑70%). Эфиры образуются либо в клетках в реакции катализируемой ацил-КоA-холестерин-ацилтрансферазой, использующей в качестве субстрата ацил‑КоA, либо в плазме в результате работы фермента лецитин-холестерин-ацилтрансферазы, осуществляющей перенос жирной кислоты со второго атома углерода фосфатидилхолина на гидроксильную группу холестерина. В плазме крови главными источниками холестерина и фосфатидилхолина для реакции служат липопротеины высокой и низкой плотности, этим путем образуется большая часть эфиров холестерина плазмы.
Для определения содержания холестерина в крови используют следующие методы:
- Титрометрические.
- Гравиметрические.
- Нефелометрические.
- Тонкослойная и газожидкостная хроматография.
- Полярографические методы, позволяют определять общий и свободный холестерин в присутствии ферментов холестеролоксидаз и холестеролэстераз.
- Флюориметрия по реакции с о‑фталевым альдегидом и другими реактивами.
- Ферментативные методы — определение протекает в одной пробирке, но в несколько этапов: ферментативный гидролиз эфиров холестерина, окисление холестерина кислородом воздуха с образованием холест‑4‑ен‑3‑ола и перекиси водорода. В качестве ферментов применяются холестеролоксидаза, холестеролэстераза, пероксидаза, каталаза. Ход реакции можно регистрировать:
- спектрофотометрически по накоплению холестенола.
- по убыли кислорода в среде.
- по изменению окраски раствора, в качестве хромогенов — индикаторов протекания реакций — используются 4‑гидроксибензоат, 4‑аминофеназон, 4‑аминоантипирин.
Все эти методы весьма специфичны и хорошо воспроизводимы.
- Колориметрические методы, в основе которых лежат следующие цветные реакции:
- реакция Биоля‑Крофта с использованием персульфата калия, уксусной и серной кислоты и с появлением красного окрашивания.
- реакция Ригли, базирующаяся на взаимодействии холестерина с реактивом, в составе которого имеются метанол и серная кислота.
- реакция Чугаева, в которой появляется красное окрашивание после реакции холестерина с ацетилхлоридом и хлористым цинком.
- реакция Либермана‑Бурхарда, при которой холестерин окисляется в сильно кислой абсолютно безводной среде с образованием сопряженных двойных связей. В результате образуется соединение холестагексаена с концентрированной серной кислотой изумрудно‑зеленого цвета с максимумом абсорбции при 410 и 610 нм. Особенностью этой реакции является отсутствие стабильности окрашивания. В литературе можно встретить разное соотношение ингредиентов в реактиве Либерман-Бурхарда: чем выше содержание уксусного ангидрида, тем с большей скоростью протекает реакция. Протеканию реакции способствуют сульфосалициловая, паратолуенсульфоновая, диметилбензол‑сульфоновая кислоты. С эфирами холестерина реакция идет медленнее, чем со свободным холестерином, скорость возрастает при повышении температуры, свет оказывает разрушающее воздействие на продукты реакции. Все методы, основанные на реакции Либерман-Бурхарда, подразделяются на прямые и непрямые:
| ◊ к непрямым относятся методы Энгельгарда‑Смирновой, Раппопорта‑Энгельберга, Абеля и заключаются в предварительной экстракции холестерина из сыворотки с последующим определением его концентрации. Из этой группы методов наиболее известен метод Абеля с экстракцией свободного и этерифицированного холестерина изопропанолом или петролейным эфиром, гидролизом эфиров холестерина и последующей реакцией Либерман-Бурхарда. Методы этой группы более воспроизводимы и специфичны; |
| ◊ в прямых методах (Илька, Мрскоса‑Товарека, Златкис‑Зака) холестерин предварительно не экстрагируется, а цветная реакция осуществляется непосредственно с сывороткой. Выявлено, что определение концентрации холестерина по Ильку при сравнении с методом Абеля дает более высокие (по разным авторам на 6%, на 10‑15%) значения, что необходимо учитывать при типировании гиперлипопротеинемий. |
- реакция Калиани‑Златкиса‑Зака, заключающаяся в появлении красно‑фиолетового окрашивания раствора при окислении холестерина хлорным железом в уксусной и концентрированной серной кислотах. Эта реакция в 4‑5 раз чувствительнее, чем реакция Либерман‑Бурхарда, но менее специфична.
Унифицированными методами являются колориметрические методы Илька и Калиани-Златкис-Зака.
Определение содержания общего холестерина в сыворотке крови методом Илька
Принцип
Основан на реакции Либерман‑Бурхарда: в сильно кислой среде в присутствии уксусного ангидрида происходит дегидратация холестерина с образованием окрашенного в зеленовато‑синий цвет бисхолестадиенилмоносульфоновой кислоты.
Нормальные величины
| Сыворотка (указанный метод) | 0 ‑ 1 год | 1.81‑4.53 ммоль/л |
| до 20 лет | 3.10‑5.80 ммоль/л | |
| 20 ‑ 29 | 3.40‑5.80 ммоль/л | |
| 30 ‑ 39 | 3.70‑6.20 ммоль/л | |
| 40 ‑ 49 | 3.70‑6.80 ммоль/л | |
| 50 ‑ 59 | 4.10‑7.30 ммоль/л | |
| 60 ‑ 69 | 4.20‑7.40 ммоль/л |
Определение количества общего холестерина в сыворотке крови методом Златкис‑Зака
Принцип
Свободный и эфирносвязанный холестерин окисляется хлорным железом в присутствии уксусной, серной и фосфорной кислот с образованием ненасыщенных продуктов, окрашенных в фиолето‑красный цвет.
Нормальные величины
| Сыворотка (указанный метод) | 3,1‑6,5 ммоль/л |
Определение содержания общего холестерина ферментативным методом по набору "Новохол"
Принцип
Основан на использовании сопряженных ферментативных реакций, катализируемых: 1) холестеролэстеразой, катализируещей гидролиз эфиров холестерина до свободного холестерина; 2) холестеролоксидазой, катализирующей превращение холестерина в холестенон с образованием перекиси водорода; 3) пероксидазой, катализирующей в присутствии фенола окисление перекисью водорода 4‑аминоантипирина с образованием окрашенного продукта розово‑малинового цвета.
Нормальные величины
| Сыворотка(указанный метод) | 20‑29 лет | 3,70‑6,51 ммоль/л |
| 30‑39 лет | 4,25‑7,04 ммоль/л | |
| 40‑49 лет | 4,37‑7,70 ммоль/л | |
| старше 50 лет | 4,55‑8,24 ммоль/л | |
| Спинномозговая жидкость | 0‑0,013 ммоль/л | |
| Слюна | 0,065‑0,233 ммоль/л | |
| Желчь | печеночная | 2,1‑5,4 ммоль/л |
| пузырная | в среднем 11,1 ммоль/л | |
Влияющие факторы
Завышение результатов при колориметрических методах исследования происходит при высоком содержании в пробе билирубина, гемоглобина, витамина A; при ферментативном методе — оксикортикостероидов и применении антикоагулянтов (фторидов, оксалатов).
Клинико‑диагностическое значение
Сыворотка
Существенное повышение содержания холестерина отмечается при гиперлипопротеинемии IIa типа (семейная гиперхолестеринемия), IIb и III типа (полигенная гиперхолестеринемия, семейная комбинированная гиперлипидемия), умеренное повышение наблюдается при гиперлипопротеинемии I, IV, V типа, а также заболеваниях печени (внутри‑ и внепеченочный холестаз), заболеваниях почек, злокачественных опухолях поджелудочной железы, гипотиреозе, заболеваниях сердечно‑сосудистой системы, беременности, сахарном диабете.
Снижение выявляется при гипертиреозе, циррозе печени, злокачественных опухолях печени, гипопротеинемии и аb‑липопротеинемии.
Спинномозговая жидкость
Накопление холестерина выявляется при менингите, опухоли или абсцессе мозга, кровоизлияниях в мозг, при рассеяном склерозе.
Снижение значений обнаруживается при церебральной и кортикальной атрофии.
Определение концентрации свободного иэтерифицированного холестерина в сыворотке крови
Свободный холестерин способен образовывать с дигитонином, томатином, пиридинсульфатом труднорастворимые соединения. Чаще всего применяют водно‑спиртовый или изопропаноловый раствор дигитонина.
Принцип
Холестерин экстрагируют из сыворотки с помощью изопропилового спирта, экстракт делят на две части, в одной определяют содержание общего холестерина. В другой порции экстракта свободный холестерин осаждают дигитонином, супернатант отбрасывают, а осадок растворяют и определяют содержание свободного холестерина любым методом. Содержание этерифицированного холестерина рассчитывают как разницу между общим и свободным.
Нормальные величины
| Сыворотка | фракция этерифицированного холестерина составляет 60‑80% от общего |
Клинико‑диагностическое значение
Коэффициент этерификации холестерина является важной функциональной пробой печени. Снижение коэффициента пропорционально снижению функции печени: острый и обострения хронического гепатита, механическая желтуха, цирроз печени. Степень этерификации также зависит от активности сывороточного фермента лецитин-холестерин-ацил-трансферазы, поэтому хранение пробы при комнатной температуре может изменить соотношение между свободной и этерифицированной фракцией холестерина.
Определение содержания α‑холестерина
Принцип
Разделение α‑ и β‑липопротеинов основано на избирательной способности липопротеинов очень низкой и низкой плотности образовывать нерастворимые комплексы с гепарином в присутствии двухвалентных катионов Mn2+. Липопротеины высокой плотности при этом остаются в надосадке, где определяют содержание α‑холестерина любым способом.
Определение α‑холестерина используется для расчета индекса атерогенности:
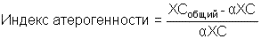
Нормальные величины
| α‑Холестерин | ||
| Сыворотка | 0,9‑1,9 ммоль/л | |
| Индекс атерогенности | ||
| Сыворотка | новорожденные | < 1.0 |
| 20-30 лет | 2.0‑2.8 | |
| больше 30 лет | 3.0‑3.5 | |
Клинико‑диагностическое значение
Возрастание концентрации α‑холестерина клинически не значимо, наблюдается при доброкачественных состояниях. Снижение содержания α‑холестерина свидетельствует об угрозе атеросклероза.
Возрастание индекса атерогенности до 4 и более наблюдается при ишемической болезни сердца и атеросклерозе.
Вы можете спросить или оставить свое мнение.
biokhimija.ru
Синтез эфиров холестерина - Справочник химика 21
Печень участвует в обмене холестерина. В ней происходит синтез эфиров холестерина, превращения холестерина с образованием холевых кислот, составных частей желчи. Печень играет важную роль в выделении холестерина из организма. Холестерин как составная часть желчи вместе с нею поступает в кишечник. [c.486] В клетках надпочечников содержание холестерина равно 10 г на 100 г ткани, т. е. в 50 раз больше средней концентрации. Основная часть холестерина в этих клетках находится в липидных каплях цитозоля, в форме эфиров. Холестерин в надпочечниках используется для синтеза стероидных гормонов. Поскольку масса надпочечников небольшая (12 г), то доля холестерина надпочечников в общем фонде невелика — примерно 1 %. [c.313]Ферментные системы печени способны катализировать все реакции или значительное большинство реакций метаболизма липидов. Совокупность этих реакций лежит в основе таких процессов, как синтез высших жирных кислот, триглицеридов, фосфолипидов, холестерина и его эфиров, а также липолиз триглицеридов, окисление жирных кислот, образование ацетоновых (кетоновых) тел и т.д. [c.556]
Источником холестерина для синтеза кортикостероидов служат его эфиры, поступающие в клетку в составе ЛНП или депонированные в клетке. Освобождение холестерина из его эфиров и синтез кортикостероидов стимулируются кортикотропином. [c.279]
Синтез и гидролиз эфиров происходят во многих клетках, но особенно активно в клетках коры надпочечников в этих клетках до 80 % всего холестерина представлено эфирами, в то время как в других клетках обычно меньше 20 %. [c.316]
Основная часть холестерина плазмы связана с липопротеидом низкой плотности (ЛНП дополнение 2-А), который доставляет эфиры холестерина непосредственно к соответствующим клеткам. Комплекс холестерин—ЛНП связывается со специфическими рецепторами ЛНП на поверхности клеток и затем захватывается клеткой путем эндоцитоза [97Ь]. Липопротеид подвергается затем действию лизосомных ферментов, а специфическая кислая липаза расщепляет эфиры холестерина и освобождает свободный холестерин. Потребление холестерина клеткой регулируется по принципу обратной связи, контролирующей интенсивность синтеза молекул рецептора ЛНП. Кроме того, свободный холестерин тормозит важный этап собственного биосинтеза — восстановление З-окси-З-метнлглутарил-СоА (рнс. 11-8) [97Ь, 97с]. Известен ряд нарушений холестеринового обмена. При одной из форм семейной гиперхоле- [c.583]
Данные о механизме действия АКТГ на синтез стероидных гормонов свидетельствуют о сугцественной роли аденилатциклазной системы. Предполагают, что АКТГ вступает во взаимодействие со специфическими рецепторами на внешней поверхности клеточной мембраны (рецепторы представлены белками в комплексе с другими молекулами, в частности с сиаловой кислотой). Сигнал затем передается на фермент аденилатцикла-зу, расположенную на внутренней поверхности клеточной мембраны, которая катализирует распад АТФ и образование цАМФ. Последний активирует протеинкиназу, которая в свою очередь с участием АТФ осуществляет фосфорилирование холинэстеразы, превращающей эфиры холестерина в свободный холестерин, который поступает в митохондрии надпочечников, где содержатся все ферменты, катализирующие превращение холестерина в кортикостероиды. [c.259]
Основная часть холестерина в зрелом мозге находится в неэтерифици-рованном состоянии, эфиры холестерина обнаруживаются в относительно высокой концентрации в участках активной миелинизации. Пути биосинтеза фосфоглицеридов в мозге сходны с теми, которые осуществляются в других тканях. Жирные кислоты образуются в основном из глюкозы, однако частично синтез их происходит из ацетоацетата, цитрата и даже ацетил-аспартата. [c.636]
Авторы синтеза получили холестерин-7-Hi также восстановлением бензоата 7-бромхолесте )ина водородом-Нз в присутствии палладиевого катализатора, нанесенного на углекислый кальций (5% Pd). После гидролиза Образующегося эфира и очистки продукта хроматографией выход холестерина-7-Hl составил 52,5%. [c.391]
Значительное число работ " посвящено бромированию эфиров холестерина. Получаемое 7-бромпроизводное может быть дегидробромировано с образованием 7-дегидрохолестерина, что является новым улучшенным синтезом провитамина Дд. [c.295]
Синтез стероидных гормонов стимулируется в Коре надпочечников кортикотропином, а в половых железах — лютропином. Эти тропные гормоны аденогипофиза регулируют транспорт эфиров холестерина в клетку активность холестеролэстеразы митохондриальные ферменты превращения холестерина в прегненолон процессы катаболизма углеводов и липидов, обеспечивающие стероидогенез энергией и пластическим материалом. [c.394]
Напишите реакции синтеза и гидролиза линолеил-холестерина, которые постоянно происходят в клетках коры надпочечников. Укажите ферменты и остальных участников этих реакций. В каких условиях в клетках преимущественно протекает образование эфиров холестерина, а в каких — его гидролиз [c.216]
ФОСФАТИДЫ (фосфолипиды) — сложные эфиры фосфорной кислоты и глицерина или сфингозина, которые связаны эфирной или амидной связью с одним или несколькими остатками высших жирных кислот. В зависимости от природы спирта, лежащего в основе химической структуры Ф., различают глицерофос-фатиды и сфингофосфатиды. Ф. входят в состав клеток и тканей всех живых организмов. Особенно велико их содержанне в нервной ткани, они есть в мозге, печени, мускулах, принимают участие в окислительных процессах живых организмов. Ф. вместе с холестерином и белками, участвуют в построении мембран клеток, обусловливают избирате,аьную проницаемость для различных соединений, активно переносят вещества через мембраны, играют важную роль в транспортировке жиров, жирных кислот и холестерина. Нарушение синтеза Ф. в организме ведет к развитию жирового перерождения печени. [c.264]
Несмотря па сложность структур описанных соединений, их химическне свойства — это преимущественно свойства простых алифатических соединений. Так, холевые кислоты образуют сложные эфиры как по карбоксильной группе, так и по спиртовой гидроксильной группе, они подвергаются окислению, давая в качестве конечных продуктов трпкетоны (через стадии обра-зовання моно- и дикетонов). Эстрадиол обладает свойствами фенола II вторичного спирта, в го время как прогестерон дает реакции, ожидаемые для простого кетона и а,р-ненасыщенного кетона (гл. 16). Холестерин ведет себя как алкен и вторичный снирт. Биологический интерес к стероидам сосредоточен на установлении взаимосвязи между структурой и физиологической активностью, а также на выяснении возможных путей синтеза этих соединений в организме. С точки зрения химии стероиды также имеют большое значение и не только сами по себе, но и из-за очень важных стереохимических закономерностей их химических реакций, которые являются в основном следствием жесткости скелета молекулы, образованного конденсированными циклами. [c.361]
Тестостерон — мелкий белый кристаллический порошок, т. пл. 166— 171°. не растворим в воде, растворяется в спирте, эфире, хлороформе. Применяют обычно в виде сложных эфиров — пропионата или энантата. При синтезе стероидных гормонов большое значение имеет доступность исходного сырья в этом отношении Р-ситостерин, получаемый из отходов производства бумаги, является весьма перспективным. Как показали исследования д. М. Халецкого с сотр., технология производства стероидных гормонов из р-ситостерина мало отличается от их производства из холестерина. Между тем, холестерин является дорогим сырьем животного происхождения, в то время как р-ситостернн — недефицитным и дешевым. [c.579]
На П стадии синтеза холестерина мевалоновая кислота превращается в сквален. Реакции П стадии начинаются с фосфорилирования мевалоновой кислоты с помощью АТФ. В результате образуется 5-фосфорный эфир, а затем 5-пирофосфорный эфир мевалоновой кислоты [c.400]
Первый пример полного синтеза стероидов был описан в 1939 г. Бахманном, осуществившим вместе с сотрудниками линейную синтетическую цепочку, при которой последовательно проводилось построение одного за другим колец А, В, С и О. Примером подобного синтеза может служить и синтез холестерина по Вудворду [3.7.3]. В последнее время ( ыли разработаны конвергентные сходящиеся) синтетические цепочки при этом сначала получают два фрагмента целевой молекулы по раздельно осуществляемым схемам, а затем, по возможности на одной из последних стадий, соединяют эти фрагменты в более сложную структуру. Такой путь синтеза позволяет уменьшить потери промежуточно синтезируемых соединений, на получение которых ступень за ступенью затрачивается много времени и средств. Поэтому конвергентные пути синтеза со сходящимися цепочками особенно привлекательны для промышленного производства стероидов. Ниже приводится пример построения стероидного скелета с помощью линейного синтеза из метилового эфира 5-оксогептен-6-овой-1 кислоты и 2-метилциклопентандиона-1,3 [3.7.4] (см. схему на с. 696). [c.695]
Ключевой продукт 1 был далее стандартными трансформациями превращен в эфир 20, послуживщий общим исходным соединением для синтеза природных стероидов 2-7. Селективное гидрирование двух сопряженных двойных связей над палладием привело к продукту 21, в котором сохранив-щаяся изолированная двойная связь в цикле В обеспечивала в дальнейшем возможность введения кислородного заместителя в положение 11. Этот путь позволил осуществить успешный синтез кортикостероидов, в том числе кортизона (7). Для получения стероидов 2—6 потребовалось полное гидрирование всех трех двойных связей С=С над платиновым катализатором, приводящее к насыщенному эфиру 22. Кроме удаления лишних функциональных групп, эта реакция приводила к созданию требуемой конфигурации хиральных центров С-10 и С-17. Поскольку превращение эфира 22 в стероиды 2—5 уже было описано ранее, выполненный Вудвордом синтез этого соединения представлял собой одновременно и завершение полного синтеза стероидов 2-5. Наконец, с помощью ряда обьршых методов (присоединение алифатического заместителя в положение 17 и трансформации функциональных групп) из эфира 22 был синтезирован холестерин (6). [c.293]
Для синтеза эпоксидных соединений обычно применяют над-бензойную кислоту, реакцию проводят в инертных растворителях (хлороформе или эфире) при низкой температуре. Мононадфталевая кислота, которая также используется для этой цели, имеет по сравнению с надбензойной то преимущество, что, во-первых, обладает большей термической устойчивостью и, во-вторых, фталевая кислота, образующаяся из нее в результате реакции, не растворима в обычно применяемых растворителях Майлс и Клиф использовали для эпоксидирования пинена и холестерина надкамфорную кислоту. [c.224]
Основное направление научных исследований — структурная органическая химия. Получил (1906) недоокись углерода. Вел работы по установлению строения холестерина и холевой кислоты, что нашло отрайназваниях кислота Дильса , углеводород Дильса , дегидрирование селеном по Диль су . Изучал совместно с К- Альде ром (1911) азодикарбоновый эфир Эти работы были прерваны в свя зи с началом первой мировой вой ны и возобновлены в 1920-е. Они послужили отправным пунктом в открытии (1928) Дильсом и Аль-дером одной из важнейших реакций современной органической химии — 1,4-присоеД11нения молекул с активированной кратной связью (диенофилов) к сопряженным диенам с образованием циклических структур (диеновый синтез). Открыл (1930) каталитическую реакцию селективного дегидрирования циклогексенового или циклогекса-нового кольца в молекулах полициклических соединений действием селена при нагревании, приводящую к образованию ароматических соединений. [c.174]
Реакция Виттига была использована в синтезах многих природных соединений, в том числе предшественника холестерина— сквалена, а также ликопина и сложных эфиров биксина. Область применения этой реакции довольно обширна. [c.65]
После завершения опытов и удаленна трития камеру для разряда откры вали и твердые продукты растворяли в спирте. Аликвоты этих растворов анализировали жидкостным сцинтилляцнопным счетчиком на определенне общего количества введенного трития. Тритий определяли но потере активности после отгонки спирта. Пальмитиновую кислоту очищали рекристаллизацией из ацетона методом хроматографии по Норнту, а во втором опыте — путем превращения ее в эфир и-бромистого фенацнла. Бензойная кислота очищалась до постоянной активности путем неоднократной рекристаллизации из воды и гексана. Исходные растворы бензойной кислоты и холестерина обесцвечивали углем. Холестерин частично очищали неоднократной рекристаллизацией из спирта и гептана. Полная очистка холестерина достигалась путем синтеза его в дибролшд и рекристаллизации этого продукта из смеси эфира и спирта. [c.93]
Из эфира З-кетоэтиоаллохолаповсй кислоты могут быть получены также и некоторые стероилныр гормоны, напримео андростерон, тестостерон и кортизон. Таким образом, рассмотренный синтез холестерина является одновремеь но и синтезом целого ряда другие стероидов. [c.178]
Синтез стероидных гормонов происходит по следующей схеме. В эндокринных клетках холестеролэстераза высвобождает холестерин из его эфиров (30—40% из них — эндогенные эфиры, 60—70% — поступает в составе ЛПНП). Свободный холестерин переносится в митохондрии, где из него образуется прегненолон, который попадает в эндоплазматическую сеть. Все дальнейшие реакции идут в эндоплазматической сети и цитоплазме с использованием метаболитов обмена углеводов и липидов. [c.394]
chem21.info