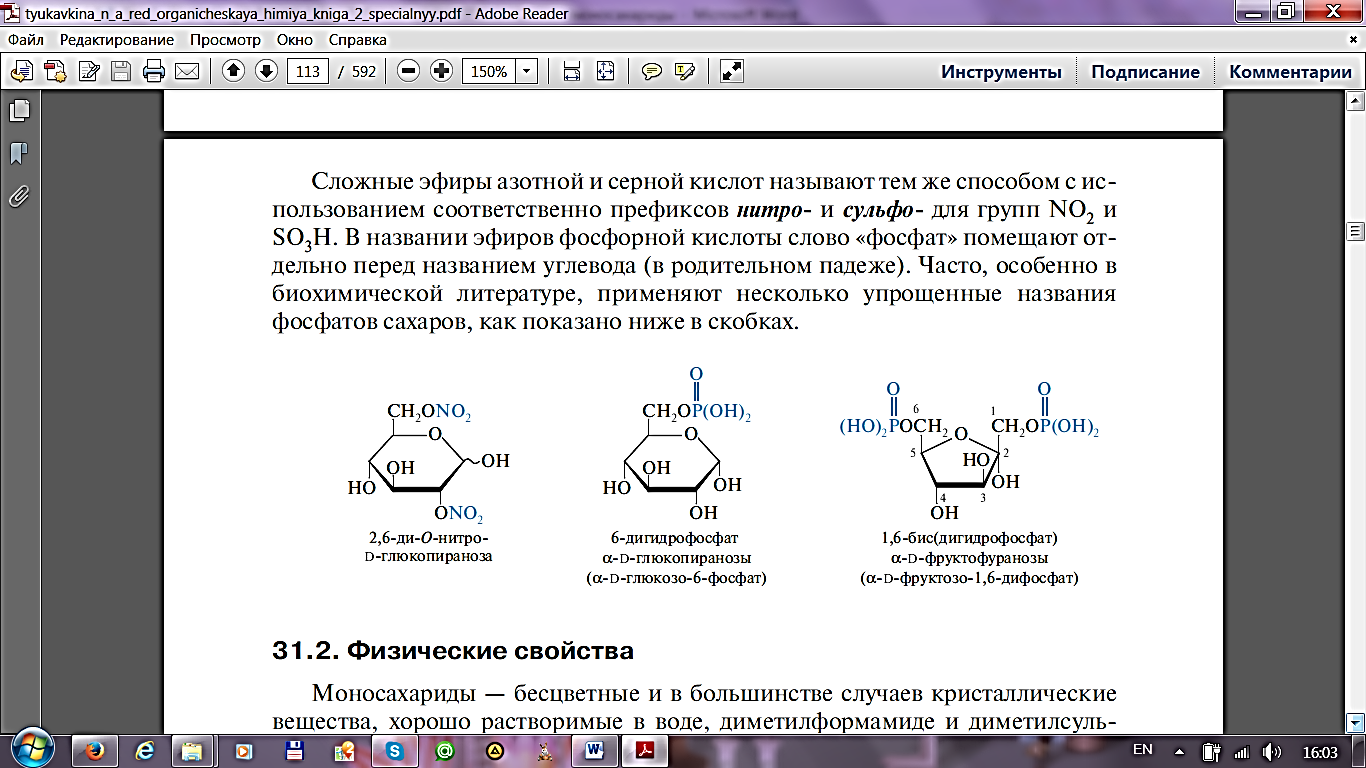
СЛОЖНЫЕ ЭФИРЫ МОНОСАХАРИДОВ. Фосфорные эфиры гексоз
Спиртовое брожение
Биохимические этапы спиртового брожения.
При спиртовом брожении помимо основных продуктов — спирта и СО2, из сахаров возникает множество других, так называемых вторичных продуктов брожения. Из 100 г С6Н12О6 образуется 48,4 г этилового спирта, 46,6 г диоксида углерода, 3,3 г глицерина, 0,5 г янтарной кислоты и 1,2 г смеси молочной кислоты, ацетальдегида, ацетоина и других органических соединений.
Наряду с этим дрожжевые клетки в период размножения и логарифмического роста потребляют из виноградного сусла аминокислоты, необходимые для построения собственных белков. При этом образуются побочные продукты брожения, главным образом высшие спирты.
В современной схеме спиртового брожения насчитывается 10—12 фаз биохимических превращений гексоз под действием комплекса ферментов дрожжей. В упрощенном виде можно выделить три этапа спиртового брожения.
I этап — фосфорилирование и распад гексоз. На этом этапе протекает несколько реакций, в результате которых гексоза превращается в триозофосфат:
|
С6Н12О6 + 2Н3РО4 Гексоза |
Фосфогексокиназа, изомериаза, альдолаза
АТФ → АДФ |
2С3Н2О3(Н2РО3) + Н2О Триозофосфат |
Главную роль в передаче энергии в биохимических реакциях играют АТФ (аденозинтрифосфат) и АДФ (аденозиндифосфат). Они входят в состав ферментов, аккумулируют большое количество энергии, необходимой для осуществления жизненных процессов, и представляют собой аденозин — составную часть нуклеиновых кислот — с остатками фосфорной кислоты. Вначале образуется адениловая кислота (монофосфат аденозина, или аденозинмонофосфат — АМФ):

Если обозначить аденозин буквой А, то строение АТФ может быть представлено в следующем виде:
ОН ОН ОН
║ ║ ║
А—О—Р—О ~ Р — О ~ Р— ОН
║ ║ ║
О О О
Значком с ~ обозначены так называемые макроэргические фосфатные связи, чрезвычайно богатые энергией, которая выделяется при отщеплении остатков фосфорной кислоты. Передача энергии с АТФ на АДФ может быть представлена следующей схемой:

Выделяющаяся энергия используется дрожжевыми клетками для обеспечения жизненных функций, в частности их размножения. Первым актом выделения энергии и является образование фосфорных эфиров гексоз — фосфорилирование их. Присоединение к гексозам остатка фосфорной кислоты от АТФ происходит под действием фермента фосфогексокиназы, поставляемого дрожжами (молекулу фосфата обозначим буквой Р):

Глюкоза Глюкозо-6-фосфат фруктозо-1,6-фосфат
Как видно из приведенной схемы, фосфорилирование происходит дважды, причем фосфорный эфир глюкозы под действием фермента изомеразы обратимо превращается в фосфорный эфир фруктозы, имеющий симметричное фурановое кольцо. Симметричное расположение остатков фосфорной кислоты по концам молекулы фруктозы облегчает ее последующий разрыв как раз в середине. Распад гексозы на две триозы катализирует фермент альдолаза; в результате распада образуется неравновесная смесь 3-фосфоглицеринового альдегида и фосфодиоксиацетона:

Фосфоглицери-новый альдегид (3,5 %) Фосфодиокси-ацетон (96,5 %)
В дальнейших реакциях участвует только 3-фосфоглицериновый альдегид, содержание которого постоянно пополняется под действием фермента изомеразы на молекулы фосфодиоксиацетона.
ІІ этап спиртового брожения — образование пировиноградной кислоты. На втором этапе триозофосфат в виде 3-фосфоглицеринового альдегида под действием окислительного фермента дегидрогеназы окисляется в фосфоглицериновую кислоту, а она при участии соответствующих ферментов (фосфоглицеромутазы и энолазы) и системы ЛДФ — АТФ превращается в пировиноградную кислоту:
|
С3Н2О3(Н2РО3) + [О] 3-Фосфоглицириновый альдегид |
Дигедрогеназа, фосфотрансфераза, фосфоглицеромутаза, энолаза |
С3Н4О3 + Н3РО4 Пировино - Фосфорная Градная кислота Кислота |
Вначале каждая молекула 3-фосфоглицеринового альдегида присоединяет к себе еще один остаток фосфорной кислоты (за счет молекулы неорганического фосфора) и образуется 1,3-дифосфоглицериновый альдегид. Затем в анаэробных условиях происходит его окисление в 1,3-дифосфоглицериновую кислоту:

Активной группой дегидрогеназы является кофермент сложного органического строения НАД (никотинамидадениндинуклеотид), фиксирующий своим никотинамидным ядром два атома водорода:
НАД+ + 2Н+ + НАД • Н2
НАД окисленный НАД восстановленный
Окисляя субстрат, кофермент НАД становится обладателем свободных ионов водорода, что придает ему высокий восстановительный потенциал. Поэтому бродящее сусло всегда характеризуется высокой восстанавливающей способностью, что имеет большое практическое значение в виноделии: понижается рН среды, восстанавливаются временно окисленные вещества, погибают патогенные микроорганизмы.
В заключительной фазе II этапа спиртового брожения фермент фосфотрансфераза дважды катализирует перенос остатка фосфорной кислоты, а фосфоглицеромутаза перемещает его от 3-го углеродного атома ко 2-му, открывая возможность ферменту энолазе образовать пировиноградную кислоту:

1,3-Дифосоглицериновая кислота 2-Фосфогглицериновая кислота Пировиноградная кислота
В связи с тем что из одной молекулы дважды фосфорилированной гексозы (израсходовано 2 АТФ) получаются две молекулы дважды фосфорилированных триоз (образовано 4 АТФ), чистым энергетическим балансом ферментативного распада сахаров является образование 2 АТФ. Эта энергия обеспечивает жизненные функции дрожжей и вызывает повышение температуры бродящей среды.
Все реакции, предшествующие образованию пировиноградной кислоты, присущи как анаэробному сбраживанию сахаров, так и дыханию простейших организмов и растений. III этап имеет отношение только к спиртовому брожению.
III этап спиртового брожения — образование этилового спирта. На заключительном этапе спиртового брожения пировиноградная кислота под действием фермента декарбоксилазы декарбоксилируется с образованием ацетальдегида и диоксида углерода, а с участием фермента алкогольдегидрогеназы и кофермента НАД-Н2 происходит восстановление ацетальдегида в этиловый спирт:

Пировиноградная кислота Ацетилальдегид Этиловый спирт
Если в бродящем сусле есть избыток свободной сернистой кислоты, то часть ацетальдегида связывается в альдегидсернистое соединение: в каждом литре сусла 100 мг Н2SO3 связывают 66 мг СН3СОН.
Впоследствии при наличии кислорода это нестойкое соединение распадается, и в виноматериале обнаруживают свободный ацетальдегид, что особенно нежелательно для шампанских и столовых виноматериалов.
В сжатом виде анаэробное превращение гексозы в этиловый спирт может быть представлено следующей схемой:

Схема спиртового брожения.
Как видно из схемы спиртового брожения, сперва образуются фосфорные эфиры гексоз. При этом молекулы глюкозы и фруктозы под действием фермента гексокеназы присоединяют остаток фосфорной кислоты от аденозиттрифосфата (АТФ), при этом образуется глюкоза-6-фосфат и аденозитдифосфат (АДФ).
Глюкоза-6-фосфат под действием фермента изомеразы превращается в фруктозу-6-фосфат, присоединяющий еще один остаток фосфорной кислоты из АТФ и образующий фруктозу-1,6-дифосфат. Эта реакция катализируется фосфофруктокиназой. Образованием этого химического соединения заканчивается первая подготовительная стадия анаэробного распада сахаров.
В результате этих реакций молекула сахара переходит в оксиформу, приобретает большую лабильность и становится более способной к ферментативным преобразованиям.
Под влиянием фермента альдолазы фруктоза-1, 6-дифосфат расщепляется на глицеринальдегидофосфорную и диоксиацетонофосфорную кислоты, способные превращаться одна в одну под действием фермента триозофосфатизомеразы. Дальнейшему преобразованию подвергается фосфоглицериновый альдегид, которого образуется приблизительно 3 % по сравнению с 97 % фосфодиоксиацетона. Фосфодиоксиацетон, по мере использования фосфоглицеринового альдегида, превращается под действием изомеразы фосфотриоз в 3-фосфоглицериновый альдегид.
На второй стадии 3-фосфоглицериновый альдегид присоединяет еще один остаток фосфорной кислоты (за счет неорганического фосфора) с образованием 1, 3-дифосфоглицеринового альдегида, который дегидруется под действием триозофосфатдегидрогеназы и дает 1, 3-дифосфоглицериновую кислоту. Водород, в этом случае, переносится на окисленную форму кофермента НАД. 1, 3-дифосфоглицериновая кислота, отдавая АДФ (под действием фермента фосфоглицераткеназы) один остаток фосфорной кислоты, превращается в 3-фосфоглицериновую кислоту, которая под действием фермента фосфоглицеромутазы превращается в 2-фосфоглицериновую кислоту. Последняя, под действием фосфопируватгидротазы, превращается в фосфоэнолпировиноградную кислоту. Дальше, при участии фермента пируваткеназы, фосфоэнолпировиноградная кислота передает остаток фосфорной кислоты молекуле АДФ, в результате чего образуется молекула АТФ и молекула энолпировиноградной кислоты переходит в пировиноградную кислоту.
Третья стадия спиртового брожения характеризуется расщеплением пировиноградной кислоты под действием фермента пируватдекарбоксилазы на диоксид углерода и уксусный альдегид, который под действием фермента алкогольдегидрогеназы (коферментом ее является НАД) восстанавливается в этиловый спирт.
Суммарное уравнение спиртового брожения может быть представлено так:
С6Н12О6 + 2Н3РО4 + 2АДФ → 2С2Н5ОН + 2СО2 + 2АТФ + 2Н2О
Таким образом, при брожении происходит преобразование одной молекулы глюкозы в две молекулы этанола и две молекулы диоксида углерода.
Но указанный ход брожения не единственный. Если, например, в субстрате нет фермента пируватдекарбоксилазы, то не происходит расщепление пировиноградной кислоты до уксусного альдегида и восстановлению подвергается непосредственно пировиноградная кислота, превращаясь в молочную кислоту в присутствии лактатдегидрогеназы.
В виноделии брожение глюкозы и фруктозы происходит в присутствии бисульфита натрия. Уксусный альдегид, образующийся при декарбоксилировании пировиноградной кислоты, удаляется в результате связывания бисульфитом. Место уксусного альдегида занимают диоксиацетонфосфат и 3-фосфоглицериновый альдегид, они получают водород от восстановленных химических соединений, образуя глицерофосфат, который превращается в результате дефосфорилирования в глицерин. Это вторая форма брожения по Нейбергу. По этой схеме спиртового брожения происходит накопление глицерина и уксусного альдегида в виде бисульфитной производной.
Вещества, образующиеся при брожении.
В настоящее время в продуктах брожения найдено около 50 высших спиртов, которые обладают разнообразными запахами и существенно влияют на аромат и букет вина. В наибольших количествах при брожении образуются изоамиловый, изобутиловый и N-пропиловый спирты. В мускатных игристых и столовых полусладких винах, получаемых путем так называемого биологического азотопонижения, в большом количестве (до 100 мг/дм3) найдены ароматические высшие спирты β-фенилэтанол (ФЭС), тирозол, терпеновый спирт фарнезол, обладающие ароматом розы, ландыша, цветов липы. Их присутствие в небольшом количестве желательно. Кроме того, при выдержке вина высшие спирты вступают в этерификацию с летучими кислотами и образуют сложные эфиры, придающие вину благоприятные эфирные тона зрелости букета.
Источником высших спиртов являются, прежде всего, аминокислоты, потребляемые дрожжами при размножении на стадии логарифмического роста.
Согласно теории Ф. Эрлиха высшие спирты образуются двумя путями:
І — через декарбоксилирование
|
 R−CH(Nh3)COOH →
R−CH(Nh3)COOH →
|
ІІ — через первоначальное гидролитическое дезаминирование
В дальнейшем было доказано, что основная масса алифатических высших спиртов образуется из пировиноградной кислоты путем переаминирования и непосредственного биосинтеза с участием аминокислот и ацетальдегида. Но наиболее ценные ароматические высшие спирты образуются только из соответствующих аминокислот ароматического ряда, например:

Образование высших спиртов в вине зависит от многих факторов. В нормальных условиях их накапливается в среднем 250 мг/дм3. При медленном длительном брожении количество высших спиртов возрастает, при повышении температуры брожения до 30 °С — уменьшается. В условиях поточного непрерывно брожения размножение дрожжей очень ограничено и высших спиртов образуется меньше, чем при периодическом способе брожения.
При уменьшении количества дрожжевых клеток в результате охлаждения, отстаивания и грубой фильтрации забродившего сусла происходит медленное накопление биомассы дрожжей и одновременно растет количество высших спиртов, прежде всего ароматического ряда.
Повышенное количество высших спиртов нежелательно для столовых белых сухих, шампанских и коньячных виноматериалов, однако придает многообразие оттенков в аромате и вкусе красным столовым, игристым и крепким винам.
Спиртовое брожение виноградного сусла связано также с образованием высокомолекулярных альдегидов и кетонов, летучих и жирных кислот и их эфиров, имеющих значение в формировании букета и вкуса вина.
Добавить комментарий
vinograd-vino.ru
| Обратная связь ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса - ваш вокал Как цель узнает о ваших желаниях прежде, чем вы начнете действовать. Как компании прогнозируют привычки и манипулируют ими Целительная привычка Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Тренинг уверенности в себе Вкуснейший "Салат из свеклы с чесноком" Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Как слышать голос Бога Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека - Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д. Отёска стен и прирубка косяков - Когда на доме не достаёт окон и дверей, красивое высокое крыльцо ещё только в воображении, приходится подниматься с улицы в дом по трапу. Дифференциальные уравнения второго порядка (модель рынка с прогнозируемыми ценами) - В простых моделях рынка спрос и предложение обычно полагают зависящими только от текущей цены на товар. |
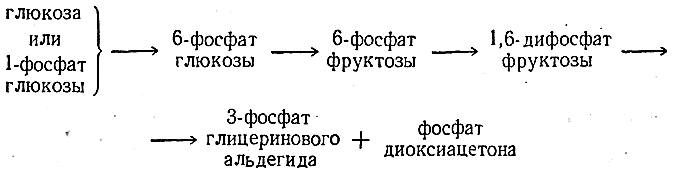
Фосфоэфиры- эфиры моносахаридов и фосфорной кислоты играют важную роль в метаболизме. Реакция фосфорилирования моносахаридов важна для метаболизма ещё и потому, что клеточная мембрана мало проницаема для этих соединений, т.е. клетка удерживает моносахариды благодаря тому, что они находятся в фосфорилированной форме.
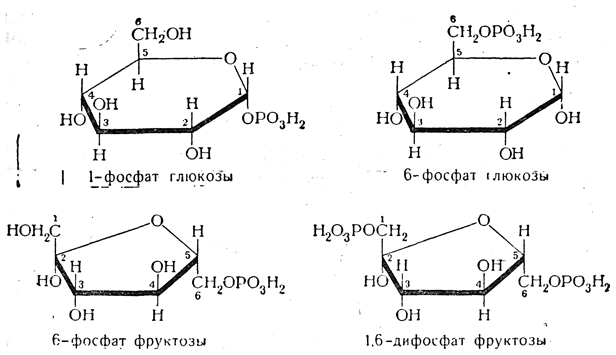
1-фосфат глюкозы 6-фосфат глюкозы 1-Фосфат глюкозы, или 1-глюкозофосфат (1-глюкозофосфорная кислота), иногда называемый «эфиром Кори», образуется из полисахаридов — крахмала и гликогена — при их расщеплении. 1-Глюкозофосфат, в свою очередь, является исходным веществом при биосинтезе крахмала и гликогена;
6-Фосфат глюкозы, или 6-глюкозофосфат (6-глюкозофосфорная кислота), метаболит гликолиза, важнейший метаболит обмена углеводов. 1-Фосфат фруктозы-метаболит гликолиза, метаболит превращения фруктозы в глюкозу.
6-Фосфат фруктозы, или6-фруктозофосфат (эфир Нейберга) (6-фруктозофосфорная кислота), Практически вся глюкоза и фруктоза, поступающие в клетку, превращаются в это соединение, что не даёт им покинуть клетку, поскольку у подавляющего большинства живых организмов нет транспортёров сахарофосфатных эфиров.
1,6-Дифосфат фруктозы, или 1,6-фруктозодифосфат (1,6-фруктозодифосфорная кислота), важнейший интермедиат гликолиза. Введенный извне обладает мощным органопротективным действием при различных патологических состояниях, связанных с дефицитом кислорода. Фруктозо-1,6-дифосфат показывает значительную протективную активность при геморрагическом и травматическом шоках и содействует успешной реанимации. Также это соединение обладает выраженным защитным эффектом на ткань головного мозга, снижая последствия гипоксического стресса. Фруктозо-2,6-бисфосфат (фруктозо-2,6-дифосфат) — регуляторная молекула всех эукариот. Усиливает гликолиз и подавляет глюконеогенез. При биологических превращениях углеводов постоянно происходит следующая цепь превращений гексозы и ее фосфорнокислых эфиров:
Галактозо-1-фосфат-участвует в синтезе глюкозы, избыток оказывает отравляющее влияние на все органы и системы организма.
Взаимопревращения фосфатов углеводов. Все поступающие с пищей моносахариды (фруктоза, галактоза, манноза и т.п.), превращаются в глюкозу через фосфорные эфиры для ее дальнейшего использования в реакциях метаболизма. Если в молекуле субстрата имеется несколько хиральных атомов углерода, то обращение конфигурации при одном из них приводит к образованию нового диастереомера. Ферменты, катализирующие такие процессы (реакции эпимеризации), называют эпимеразами. 1)Эпимеризация рибулозо-5-фосфата:
Причем известны ферменты, катализирующие эпимеризацию не только углеводов, но и их нуклеозиддифосфатпроизводных. Например, ключевым процессом в образовании и деструкции галактозы – компонента молочного сахара лактозы – является эпимеризация уридиндифосфатглюкозы (УДФ-глюкозы). 2). Эпимеризация УДФ-глюкозы:
3). Изомеризация глюкозо-6-фосфата в фруктозо-6-фосфат:
Еще одним примером является взаимопревращение изомерных глюкозо-1-фосфата и глюкозо-6-фосфата. Установлено, что взаимопревращение этих глюкозо-монофосфатов происходит только в присутствии глюкозо-1,6-дифосфата. 4). Взаимопревращение глюкозо-1-фосфата и глюкозо-6-фосфата:
Играют значительную роль в патологических процессах: воспаления, иммунного ответа, канцерогенеза (некоторые из сиаловых кислот, например, N-ацетилнейраминовая кислота участвует в проникновении метастазов в кровеносные сосуды), проникновения вирусов, бактерий и грибков в организм человека и тд.)
|





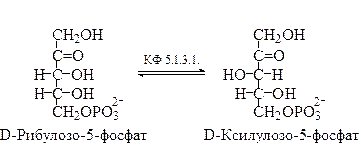


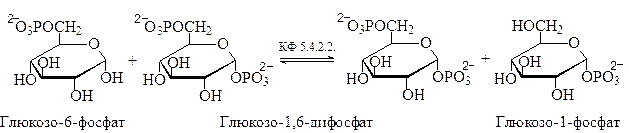

megapredmet.ru
Фосфорнокислые эфиры - Справочник химика 21
В виде своего фосфорнокислого эфира холин входит в состав веществ класса липидов, а именно фосфатидов — лецитинов (стр. 331)- Лецитины построены как смешанные глицериды жирных кислот и фосфата холина, т. е. одной из их возможных формул является формула типа [c.365]ФОСФОРНОКИСЛЫЕ ЭФИРЫ МОНОСАХАРИДОВ 661 [c.661]
Температура проведения процесса Hii должна быть слишком низкой, так как уже при 130°С вместо полимеризации идет образование фосфорнокислых эфиров. Нельзя допускать также чрезмерного повышения температуры, так ка выше 220°С увеличивается вероятность распада полимерных карбкатионов. Кроме того, при высокой температуре интенсифицируется реакция отрыва гидрид-иона от исходного алкена, в результате чего увеличивается выход смолообразных ненасыщенных продуктов, блокирующих поверхность катализатора. Выше 270°С пс лимеризация становится термодинамически невозможной. [c.267]
Нуклеотиды относятся к фосфорнокислым эфирам их молекулы построены по схеме [c.182]Если К бродильной жидкости прибавить фтористый натрий, то можно прервать брожение на этой стадии и выделить из жидкости глицерин и глицериновую кислоту в виде фосфорнокислых эфиров. [c.337]
Одним из характерных свойств фосфорнокислых эфиров является склонность к легкой миграции в них фосфорильной группировки к соседней свободной гидроксильной группе, особенно в кислой среде. Этот обмен легко происходит в том случае, когда соседние гидроксилы находятся в цис-положении, и, как было доказано, протекает через образование промежуточного циклического эфира. [c.79]
Крахмал является одним из важнейших продуктов фотосинтеза, образующимся в зеленых частях растений в виде так называемых первичных зерен. Крахмал в зеленых частях растения расщепляется на моносахариды или их фосфорнокислые эфиры и переносится в другие части растений, например в клубни картофеля или зерна злаков. Здесь происходит отложение крахмала в виде зерен. Крахмальные зерна различных растений имеют неодинаковую величину, форму и строение, так что по виду крахмальных зерен можно определить их происхождение. Наиболее высоко содержание крахмала в зернах риса (62—82%), кукурузы (65—72%), пшеницы (57—75%) и клубнях картофеля (12—24%), из которых обычно и получают крах- мал в производственной практике. [c.701]
В растениях широко распространен фитин, который представляет собой фосфорнокислый эфир мезоинозита [c.554]
На этой стадии оба соединения могут быть выделены в виде фосфорнокислых эфиров. Затем глицериновая кислота окисляется до пировиноградной кислоты [c.643]
Фосфорнокислые эфиры фенолов дают при нагревании с цианистым калием нитрилы [c.42]
Фосфорнокислые эфиры моносахаридов [c.661]
К фосфорнокислым эфирам, имеющим весьма большое зна чение, относятся следующие [c.661]
При биологических превращениях углеводов (гликолизе и спиртовом брожении) постоянно происходит следующая цепь превращений гексозы и ее фосфорнокислых эфиров [c.662]
ФОСФОРНОКИСЛЫЕ ЭФИРЫ МОНОСАХАРИДОВ [c.663]
Фосфорилирование, включая также превращение фенолов в фосфорнокислые эфиры происходит при действии галогенидов фосфора на фенолы [c.366]
Сочетание сахара (рибозы или дезоксирибозы) с органическим основанием (аденин, гуанин, цитозин или урацил) называется ну-клеозидом. Нуклеотиды являются фосфорнокислыми эфирами нуклеозидов. [c.182]
Затем происходит расщепление фосфорнокислых эфиров на диоксиацетон и глицериновый альдегид, находящиеся в динамическом (таутомерком) равновесии [c.337]
Зерно крахмала построено из двух различных веществ ами-лопектина, составляющего оболочку зерна, и амилозы—внутренней части зерна количество амилопектина относится к количеству амилозы примерно как 2 1. Амилопектин дает клейстер амилоза образует настоящий коллоидный раствор, не проявляющий никаких свойств клейстера. Амилоза окрашивается иодом в чисто синий цвет, амилопектин—в красновато-фиолетовый. Амилоза—углевод, амилопектин—фосфорнокислый эфир другого углевода. Молекулы амилозы имеют линейное строение, а молекулы амилопектина—сильно разветвлены. [c.345]
В зависимости от участия ферментов трансальдолазы или транскетолазы из двух остатков триоз возникает фруктоза или глюкоза в виде соответствующих фосфорнокислых эфиров. Этот процесс обратим и в живой клетке древесины из )-фруктозо-6-фосфата с помощью ферментов может образоваться глицеротрифосфат. [c.332]
Впервые пигмент, содержащий флавин, выделен в 1879 г. из сыворотки коровьего молока [129]. Затем из молока был выделен желтый водорастворимый фактор [130, 131], названный лактохромом [131], он же был выделен из одуванчика, солода [132], яичного желтка [133], печени [133, 1341 из свиного сердца вьщелено желтое вещество — цитофлав (оказавшееся фосфорнокислым эфиром рибофлавина) [1351. Варбург и Христиан [22] в 1932 г. выделили из дрожжей желтый окислительный фермент, в котором ими был открыт рибофлавин (лактофлавин), являющийся составной частью этого фермента. [c.522]
Сложные эфиры различных тоноалкильных эфиров этиленгликоля и некоторых сульфированных карбоновых кислот (например сульфированной рицинолевой кислоты) рекомендовались как смачивающие, ттенообразующие и диспергирующие средства Полные фосфорнокислые эфиры моноалкил- или моноарил-эфиров этилен-, пропилен- и бутиленгликолей можно приготовить действием хлорокиси фосфора на моноэфирьг. Эти соединения предлагались как пластификаторы для нитроцеллюлозных лаковых композиций. Они обладают хорошей устойчивостью к действию света и к низким температурам [c.570]
Фитин (фосфорнокислый эфир мезо-инозита) обладает активностью для мышей, но неактивен для микроорганизмов, по-видимому, вследствие их неспособности расщеплять связанную форму инозита. Алкильные эфиры лезо-инозита активностью для микроорганизмов также не обладают [431. Причина благоприятного действия инозита в различных случаях состоит в том, что, являясь ростовым фактором для некоторых микроорганизмов кишечной флоры, он стимулирует микробиологический синтез недостающих витаминов (например, биотина) (451. [c.625]
Еще одно простое пиридиновое основание, относящееся к витаминам, именуется пиридоксалем и имеет химическое строение 6.136, Альдегид 6.136 и его фосфорнокислый эфир повсеместно распространены в природе. Они синтезируются бактериями, растениями и грибами, входят в состав большого числа жизненно важных ферментов, катализирующих химические превращения аминокислот и аминов. Животные должны получать пиридоксаль с пи- [c.458]
Кроме свободной формы, рибофлавин встречается большей частью в виде рибофлавин-5 -фосфорнокислого эфира (флавинмононуклеотцд) и фермента d-аминокислого оксида (флавжнадениндинуклеотид). Рибофлавин трудно растворим в воде. Для количественного определения его можно экстрагировать, например, смесью пиридин — ледяная уксусная кислота — вода (10 + 1 + 40). [c.240]
Представляет интерес способ огнезащиты тканей, основанный на образовании фосфорнокислых эфиров целлюлозы. Этот способ состоит в обработке ткани при повышенной температуре смесью фосфорной кислоты с азотсодержащими основаниями мочевиной, дицианди-амидом, меламином и др. [c.19]
Установлено, что чрезвычайно важную роль в процессе брожения играют производные фосфорной кислоты. Первой стадией процесса являетсй образование из углевода (гексозы) сложных эфиров фосфорной кислоты. Источником фосфорной кислоты является аденозинтрифосфорная кислота (в биохимической литературе часто называемая сокращенно АТФ), отдающая один из трех своих остатков фосфорной кислоты и превращающаяся в аденозинднфосфорную кислоту (АДФ). При этом сначала образуются монофосфаты, а затем дифосфат гексозы—1,6-фруктозодифосфат (см. стр. 662). Открытая форма такого эфира расщепляется при помощи ферментА альдолазы на молекулу фосфорнокислого эфира глицеринового альдегида и на молекулу фосфорнокислого эфира диоксиацетона, которые могут изомеризоваться друг в друга. [c.216]
Фосфорнокислый эфир глицеринового альдегида связывает молекупу фосфорной кислоты, превращаясь в 1,3-дифосфат гидратной формы глицеринового альдегида, который дегидрируется с образованием 1,3 дифосфата глицериновой кислоты. Присоединение второй молекулы фосфорной кислоты к фосфату глицеринового альдегида, по-видимому, необходимо для практически одновременно идущего дегидрирования. Образовавшийся 1,3-дифосфат глицериновой кислоты, отдавая остаток фосфорной кислоты, находившийся при карбоксильной группе, переходит в 3-фосфат глицериновой кислоты. Далее 3-фос-фат под действием фермента фосфоглицеромутазы изомеризуется в 2-фосфат глицериновой кислоты, который действием фермента енолазы дегидратируется и превращается в фосфат енольной формы пировиноградной кислоты, о соединение, теряя фосфатный остаток, превращается в пировиноградную кислоту, дающую уксусный альдегид и СО,. Уксусный альдегид подвергается ферментативному гидрированию и превращается в этиловый спирт. [c.216]
Уже давно было обращено внимание на то, что in vitro (лат.—в стекле) сахара обладают значительной стойкостью, тогда как в организмах—in vivo-(лат.—в живом) чрезвычайно быстро идут как процессы расщепления моносахаридов (брожение, окисление), так и синтетические процессы (например, образование крахмала, гликогена). Когда Э. Фишер получил 7-метилглюкозид и оказалось, что он гидролизуется разбавленными кислотами почти в 100 раз-быстрее, чем обычные а- и р-глюкозиды, возникла идея, что в организмах глюкоза при нормальных условиях переходит в особую активную форму. Такую-неизвестную активную форму стали называть у-сахаром, не связывая сначала с этим названием какого-либо представления о структуре. В дальнейшем, когд у 7-глюкозидов было доказано наличие пятичленного кольца, у-сахарами стали называть фуранозы. Однако впоследствии оказалось, что наибольшей реакционной способностью обладают не фуранозы, а оксо-формы. Кроме того, было обнаружено, что при углеводном обмене в организмах простые сахара, прежде-чем расщепиться, как правило, превращаются в фосфорнокислые эфиры (см. стр. 662). [c.639]
Кетоза, соответствующая рибозе, носит название рибулозы. В виде фосфорнокислых эфиров она принимает участие в фотосинтезе (см. стр. 662). [c.655]
Фосфорнокислые эфиры глюкозы и фруктозы были выделены как из животных, так и из растительных 01рганизмов. Оказалось, что эти эфиры играют исключительно важную роль в жизнедеятельности организмов, являясь промежуточными продуктами расщепления углеводо в при брожении и гликолизе (расщеплении полисахарида гликогена до молочной кислоты, происходящем во всех клетках животных организмов), а также при син-тетичеоких процессах (например, при фотосинтезе). [c.661]
Дифосфат фруктозы, или 1,6-фруктозодифосфат (1,6-фруктозодифос-форная кислота), был открыт почти одновременно Л. А. Ивановым и Харденом и Юнгом. Этот эфир образуется в животных и растительных тканях путем фосфорилирования 6-фосфата фруктозы аденозинтрифосфорной кислотой. 1,6-Дифосфат фруктозы является наиболее лабильным из описанных фосфорнокислых эфиров гексоз, так как и в биологических условиях и при действии разбавленных шелочей он легко расщепляется с образованием молекул с тремя углеродными атомами. 1,6-Фрукюзодифосфорная кислота имеет восстанавливающие свойства. Ее удельное вращение [а]д = -ЬЗ,4° [c.662]
chem21.info
Взаимопревращения моносахаридов
Конечный продукт цикла Кальвина фруктозо-6-фосфат путём изомеризации очень легко превращается в глюкозо-6-фосфат под действием фермента глюкозофосфатизомеразы:
фруктозо-6-фосфат глюкозо-6-фосфат
Вследствие того, что реакции изомеризации сопровождаются небольшим изменением свободной энергии, они в большинстве своём обратимы.
Под действием фермента маннозофосфатизомеразы фруктозо-6-фосфат изомеризуется в маннозо-6-фосфат:
фрукто-6-фосфат маннозо-6-фосфат
Врезультате действия соответствующих мутаз, осуществляющих внутримолекулярный перенос фосфатной группы, из глюкозо-6-фосфата образуется глюкозо-1-фосфат, а из маннозо-6-фосфата – маннозо-1-фосфат:
глюкозо-6-фосфат глюкозо-1-фосфат
маннозо-6-фосфат маннозо-1-фосфат
В нефотосинтезирующих клетках растений и у гетеротрофных организмов фруктозо-6-фосфат и глюкозо-6-фосфат включаются в реакции гликолиза или пентозофосфатного цикла. В ходе реакций гликолиза фосфорилированная гексоза расщепляется на две триозы – 3-фосфогли-цериновый альдегид и фосфодиоксиацетон, из которых при обращении реакций гликолиза может снова синтезироваться гексоза. В пентозофосфатном цикле образуется примерно такой же набор фосфорнокислых производных моносахаридов, как и в цикле Кальвина. Таким образом, мы видим, что в гликолитических реакциях осуществляются взаимные превращения гексоз и триоз на уровне их фосфорнокислых эфиров, а в пентозофосфатном цикле и цикле Кальвина происходят взаимные превращения фосфорилированных форм триоз, пентоз, эритрозы, глюкозы и фруктозы.
Включение свободных моносахаридов в различные реакции их превращений осуществляется путем фосфорилирования. В качестве донора фосфатных групп для реакций фосфорилирования чаще всего используется АТФ. Ферменты, катализирующие перенос фосфатных групп от АТФ на моносахариды, как мы уже знаем, называют киназами.
Фосфорилирование глюкозы и маннозы катализирует фермент гексокиназа с образованием глюкозо-6-фосфата и маннозо-6-фосфата.
По аналогичному механизму фруктокиназа катализирует образование фруктозо-1-фосфата из фруктозы, а фермент галактокиназа синтезирует галактозо-1-фосфат из галактозы.
Образование свободных моносахаридов из их фосфорилированных форм катализируют ферменты гидролитического типа – фосфатазы, которые отщепляют от фосфатов моносахаридов остатки фосфорной кислоты. Гидролиз глюкозо-6-фосфата происходит с участием глюкозо-6-фосфатазы, галактозо-1-фосфата – под действием галактозо-1-фосфатазы, фруктозо-6-фосфата – фруктозо-6-фосфатазы и т.д.
глюкозо-6-фосфат глюкоза
Значительно сложнее происходят взаимопревращения галактозы и других моносахаридов. В этих реакциях в качестве промежуточных продуктов образуются нуклеозиддифосфатпроизводные соответсвующих моносаха-ридов. На первом этапе таких превращений галактоза фосфорилируется с участием фермента галактокиназы, в результате образуется галактозо-1-фосфат:
галактоза галактозо-1-фосфат
На следующей стадии галактозо-1-фосфат взаимодействует с уридинтрифосфатом (УТФ). Эту реакцию катализирует ферментгалактозо-1-фосфатуридилилтрансфераза, под действием которого обра-
галактозо-1-фосфат УДФ-галактоза
зуется нуклеотидное производное галактозы – уридиндифосфат-галактоза (УДФ-галактоза) и пирофосфат.
В дальнейшем УДФ-галактоза изомеризуется в УДФ-глюкозу под действием специфической НАД-зависимой 4-эпимеразы:
УДФ-галактоза УДФ-глюкоза
После гидролитического расщепления УДФ-глюкоза распадается на два продукта – глюкозо-1-фосфат и уридинмонофосфат (УМФ):
УДФ-глюкоза глюкозо-1-фосфат
Глюкозо-1-фосфат может далее изомеризоваться в глюкозо-6-фосфат, а глюкозо-6-фосфат – во фруктозо-6-фосфат. Таким образом, посредством указанных реакций галактоза может превращаться во фруктозо-6-фосфат, который включается в реакции дыхания, или в глюкозо-6-фосфат, способный превращаться в продукты пентозофосфатного цикла.
Возможен также синтез галактозо-1-фосфата из глюкозо-1-фосфата, так как в клетках организмов содержится фермент глюкозо-1-фосфатури-дилилтрансфераза, катализирующий образование УДФ-глюкозы из глюкозо-1-фосфата и УТФ:
глюкозо-1-фосфат + УТФ ¾¾®УДФ-глюкоза + Н4Р2О7
Затем УДФ-глюкоза под действием 4-эпимеразы изомеризуется в УДФ-галактозу, при гидролизе которой образуется галактозо-1-фосфат:
УДФ-галактоза + Н2О¾¾®галактозо-1-фосфат + УМФ
Взаимопревращения гексоз и пентоз осуществляются в пентозофосфатном цикле и цикле Кальвина. Важное значение для этих реакций имеют ферменты транскетолаза и трансальдолаза, а в пентозо-фосфатном цикле – ещё и фермент фосфоглюконатдегидрогеназа, ката-лизирующий окислительное декарбоксилирование 6-фосфоглюконовой кислоты с образованием рибулозо-5-фосфата. Этот фермент фактически осуществляет превращение гексозы в пентозу. Во взаимных превращениях пентоз также участвуют ферменты рибулозо-фосфатэпимераза и рибозофосфатизомераза, поддерживающие динамическое равновесие между рибулозо-5-фосфатом, с одной стороны, и ксилулозо-5-фосфатом и рибозо-5-фосфатом, с другой стороны.
Ксилоза и арабиноза синтезируются также из гексоз, но другим путём. При этом в качестве промежуточных продуктов образуются нуклеотидные производные глюкуроновой и галактуроновой кислот. На первом этапе осуществляется ситез УДФ-глюкозы из глюкозо-1-фосфата и
УТФ, а затем под действием фермента УДФ-глюкозодегидрогеназы(1.1.1.22) УДФ-глюкоза окисляется в УДФ-глюкуроновую кислоту:
УДФ-глюкоза УДФ-глюкуроновая
кислота
Затем УДФ-глюкуроновая кислота подвергается декарбоксили-рованию и превращению в пиранозную форму УДФ-ксилозы:
УДФ-глюкуроновая УДФ-ксилоза
кислота
Полученная таким путём УДФ-ксилоза используется в процессе синтеза ксиланов.
По аналогичному механизму осуществляется синтез УДФ-арабинозы из УДФ-галактозы, при этом в качестве промежуточного продукта образуется УДФ-галактуроновая кислота. УДФ-арабиноза так же, как и УДФ-ксилоза, не накапливается в растительных тканях, а используется для синтеза арабанов. Кроме того, возможны взаимные превращения УДФ-глюкуроновой и УДФ-галактуроновой кислот, а также пираназных форм УДФ-ксилозы и УДФ-арабинозы под действием соответствующих 4-эпимераз.
УДФ-глюкуроновая кислота УДФ-галактуроновая кислота
УДФ-галактуроновая кислота является основным источником галактуроновой кислоты для синтеза пектиновых веществ, а УДФ-глюкуроновая кислота участвует в синтезе ксиланов (в качестве ответвлений), полиуренидов, аскорбиновой кислоты.
Превращение моносахаридов в спирты. У растений, грибов и водорослей важную роль в углеводном обмене играют спирты – сорбит, маннит, дульцит.
Дульцита много содержится в листьях ряда растений, особенно в листьях бересклета. Сорбита много накапливается в плодах и ягодах. Очень много маннита в грибах и водорослях, а также в некоторых растительных продуктах. Синтез этих спиртов осуществляется в результате восстановления соответствующих моносахаридов. Наиболее хорошо изучена реакция образования маннита из фруктозы. Эту реакцию катализирует фермент маннитолдегидрогеназа (1.1.1.138):
СН2ОН СН2ОН
| |
C=OНO–C–H
| |
НO–C–H+ НАДФ×Н + Н+¾®НO–C–H+ НАДФ+
| |
H–C–OH H–C–OH
| |
H–C–OH H–C–OH
| |
Ch3OH Ch3OH
фруктоза маннит
Спирты, синтезируемые в результате восстановления моносахаридов, выполняют функцию резервных углеводов. Они очень легко превращаются в соответствующие моносахариды.
studfiles.net
Презентация на тему: Моносахариды (гексозы)
альдегидоспирт | кетоноспирт | альдегидоспирт |
Олигосахариды
Глюкоза + Глюкоза = Мальтоза Глюкоза + Галактоза = Лактоза Глюкоза + Фруктоза = Сахароза
Олигосахариды
Образование 1,4-гликозиднойсвязи между моносахаридами
М о н о с а х а р и д |
|
|
| М о н о с а х а р и д | ||||||
| 6 |
| О |
|
|
|
| 6 | О |
|
5 |
|
|
|
|
|
| 5 |
| ||
4 |
|
| 1 | О | Н | Н | О | 4 |
| 1 |
О Н | 3 |
| 2 |
|
|
|
| 3 | 2 | Н |
- Н | 2 О |
|
|
|
|
|
| + Н | 2 О |
|
К о н д е н |
|
|
|
|
|
| Г и д р о л и з |
| ||
Н 2 | О |
| Д | и | с | а х а | р | и д |
|
|
| 6 |
|
|
| ||||||
|
| О |
|
|
|
| 6 | О |
| |
| 5 |
|
|
|
| 5 |
|
| ||
4 |
|
| 1 |
| О | 4 |
| 1 |
| |
О | Н | 3 | 2 |
|
| Н | ||||
|
|
|
| 3 | 2 | |||||
1 , 4 - г л и к о з и д н а я с в я з ь
Олигосахариды
Образование 1,4-гликозиднойсвязи между моносахаридами
ПОЛИСАХАРИДЫ:
- Гомополисахариды - Гетерополисахариды
Цепи
полисахаридов
Строение, свойства, биологическая роль гомо- и гетерополисахаридов 29
studfiles.net
Монофосфат гексозы - Справочник химика 21
Дальнейшие энзиматические превращения фосфоглицериновой кислоты ведут к образованию карбоновых кислот, углеводов и аминокислот. Синтез гексоз проходит в обратном направлении, тем же путем, как гликоли т. е. энзиматический распад углеводов с участием процессов фосфоролиза,. наблюдаемый при спиртовом брожении, в мышечных тканях и т. д. Предшественниками сахарозы являются не глюкоза и фруктоза, а продукты их фосфорилирования, а именно глюкозо-1-монофосфат (эфир Кори) и фрук-тозо-6-монофосфат. Это видно из того, что в сахарозе и в обоих эфирах, при кратковременном освещении, радиоактивный углерод появляется раньше, чем в глюкозе и фруктозе. Образование аминокислот, из которых синтезируются белки, идет в общих чертах следующим путем. Двууглеродные группы типа ацетатов образуют пировиноградную и щавелеуксусную кислоты, аминирование которых дает аминокислоты. Синтез последних из ацетатов был подтвержден прямым путем добавление к освещаемой взвеси хлореллы ацетата, меченного радиоактивным С в карбоксиле, быстро ведет к появлению радиоактивного а-аланина с меченым углеродом не только в карбоксиле, но и в углеродной цепи. Превращения пировиноградной кислоты по рассматриваемому ниже циклу трикарбоновых киСт лот, повидимому, при фотосинтезе не происходит, так как не удалось идентифицировать образования радиоактивной а-кетоглютаровой кислоты и некоторых других звеньев этого цикла. Во всех рассмотренных превращениях принимают участие энзимы и процессы фосфорилирования и дефое-форилирования, как и в других случаях обмена углеводов. [c.309] Установлено, что чрезвычайно важную роль в процессе брожения играют производные фосфорной кислоты. Первой стадией процесса является образование из углевода (гексозы) сложных эфиров фосфорной кислоты. Источником фосфорной кислоты является аденозинтрифосфорная кислота (в биохимической литературе часто называемая сокращенно АТФ), отдающая один из трех своих остатков фосфорной кислоты и превращающаяся в аденозиндифосфорную кислоту (АДФ). При этом сначала образуются монофосфаты, а затем дифосфат гексозы, производное фрукто-фуранозы (см. стр. 571). Открытая форма такого эфира расщепляется при помощи фермента альдолазы на молекулу фосфорнокислого эфира глицеринового альдегида и на молекулу фосфорнокислого эфира диоксиацетона, которые могут изомеризоваться друг в друга. [c.207]Если бы образование гексозофосфатов происходило путем обращения гликолитического пути от ФГК, то углеродные атомы гексозы в положениях 3 и 4 должны были бы образоваться из атома углерода карбоксильной группы ФГК углеродные атомы в положениях 2 и 5 должны были бы происходить из а-углеродных атомов ФГК, а углеродные атомы 1 и 6 — из р-углеродных атомов ФГК. Когда Кальвин и сотр. [19] расщепили молекулы гексозы таким образом, чтобы получить названные пары углеродных атомов, они нашли, что распределение радиоактивного углерода в гексозе было именно таким, как это и предполагалось, исходя из постулированного механизма (см. фиг. 217). Кальвин и его сотрудники пришли к заключению, что ФГК, образовавшаяся в ходе фотосинтеза в результате первичной реакции карбоксилирования, восстанавливается в глицераль-дегидфосфат (V) [уравнения (10) и (11)], который изомеризуется в диоксиацетонфосфат (VI). Эти два триозофосфата затем конденсируются, образуя фруктозо-1,6-дифосфат (VII) и далее, в конечном счете, монофосфаты фруктозы и глюкозы. [c.540]
Распределение радиоактивности в монофосфатах гексоз, образующихся в этих же условиях (в %) [c.582]
Смит пришел к заключению, что первичным продуктом фотосинтеза является общий предшественник сахарозы и крахмала (быть может, монофосфат гексозы), а свободные моносахариды, найденные в клеточном соке, — это вторичные продукты, получившиеся при гидролизе сахарозы. [c.50]
Карбоксилирование рибулозодифосфата начинает восстановительный пентозофосфатный цикл углерода, звенья которого и их взаимосвязь были установлены, в частности, с помощью анализа продуктов ассимиляции СОг. Нестойкая р-кетокислота распадается при действии воды до 3-кислого фосфорнокислого эфира глицериновой кислоты. Последняя является ключевым соединением в цепи ферментативного синтеза углеводов, карбоновых кислот, аминокислот. В синтезе углеводов именно это соединение служит центром аккумулирования энергии, запасенной в ассимиляционном факторе. Сначала она фосфорилируется по карбоксильной группе с помощью АТФ, затем сложноэфирная группа триозы восстанавливается НАДФН до альдегидной, и полученный кислый фосфорнокислый эфир глицеринового альдегида превращается далее уже без участия ассимиляционного фактора АТФ принимает участие еще только в одной стадии превращения углеводов, а именно в процессе синтеза рибулозодифосфата из монофосфата. Регенерация рибулозодифосфата замыкает цикл. Все последующие изображенные на схеме превращения (см. схему 1) не требуют подвода энергии извне. Баланс по углероду показывает, что для построения одной новой молекулы гексозы требуется б актов [c.35]
Поэтому следует ожидать, что некоторые фенамины могут действовать как переносчики кислорода в процессах метаболизма биологических систем.В идеа-лефеназин должен быть тем веществом, которое образует наполовину восстановленный феназин в полном равновесии с компонентами клетки и воздухом, восстанавливаясь в процессе метаболизма клетки и быстро окислясь сноваф воздухом. Как видно из противоречивых сообщений, это условие соблюдается редко. Диккенс и Мак-Илвейн [151] показали, что феназины могут действовать как переносчики кислорода в метаболизме монофосфатов гексоз некоторыми тканями и что их активность может соответствовать в первом приближении нормальным потенциалам, как показано в табл. 6. [c.542]
В опыте с монофо офатом раствор после опыта содержит лишь следы неорганического радиоактивного фосфора., В экстракте, выделенном из корня, основная масса фосфатов была в виде монофосфата гексозы. Дифосфат гексозы при постунлен] неорганического фосфата образовывался в 1корнях лишь в незначительных количествах, а при поступлении. монофосфата гексозы — в значительном количестве. Этот факт также исключает предположение, что обнаруживаемый монофосфат гексозы может быть лишь монофосфатом, прочно адсорбированным поверхностью Корней. [c.60]
Если второго фосфорилирования гексозо-6-монофосфата не происходит, то фосфорилированная глюкоза может подвергаться прямому окислению до фосфопентоз. В норме доля пентозофосфатного пути в количественном превращении глюкозы обычно невелика, варьирует у разных организмов и зависит от типа ткани и ее функционального состояния. [c.353]
Для проверки этого иредположения были поставлены опыты с монофосфатом и дифосфатом гексозы. Меченный по фосфору глюкозо-1-фосфат был получен 1МСТ0Д0М Напез а с помощью картофельной фосфори-лазы, а 1,6-дифосфат фруктозы — при -брожении пивных дрожжей. Полученные меченые соединения очищались двукратным хроматографированием. [c.59]
Хроматограмма опыта 2 с дифосфатом гексозы также подтверждает поступление в корни фосфорорганического С0един1ени я. Поступившие в корень фосфорорганические соединения, так же как и неорганический фосфат, вступают в обменные реакции, одинаково усваиваясь растением, о чем говорит почти полная идентичность хроматограмм экстрактов листьев во всех трех опытах. В листьях содержится большое количество неопганнчеокого фосфора, а из ор-ганического присутствуют нуклеотиды, дифосфат И монофосфаты тексоз, фосфоглицериновая кислота, глицерофосфат и триозофосфат. [c.60]
Дифосфаты -Гексозо-монофосфат>1 [c.86]
Рис, 7,17. Возрастание концентрации ФГК и гексозо-монофосфатов (ГМФ) в строме хлоропластов во время индукции [26]. (В этом опыте и на рис. 7.16 длительность индукции составляла около 3 мин.) ПМФ — пен-тозомонофосфатщ. [c.184]
chem21.info
Презентация на тему: Химические свойства:
1.Углеводы обладают свойствами восстановителей (благодаря наличию
альдегидной группы в составе их молекулы), что даёт возможность проводить качественное и количественное определение сахаров.
2.При окислении моносахаридов образуются уроновые кислоты, из
которых важнейшей является глюкуроновая кислота, входящая в состав основного вещества соединительной ткани
3.Моносахариды способны образовывать эфиры, особо важны фосфорные эфиры гексоз (глюкозы, фруктозы, галактозы) и пентоз (рибозы и дезоксирибозы), так как именно фосфорилированные сахара участвуют в реакциях метаболизма.
4. Моносахариды могут присоединять аминогруппу (образуются глюкозамины) и ацетилироваться.
Моносахариды связываются друг с другом.
Мальтоза α(1→4) гликозидная связь
| Ch3OH | O |
|
| Ch3OH |
|
H | H | H | H | H | O OH | |
| OH | H | 1 | O | OH | H |
HO |
|
|
| H | ||
|
|
|
|
| ||
| H | OH |
|
| H | OH |
Ферменты обладают специфичностью по отношению к типу гликозидной связи, что имеет важнейшее значение в питании. Так,
амилаза, расщепляющая крахмал и гликоген, является α- гликозидазой. Фермент,
расщепляющий β-гликозидныесвязи, у человека отсутствует, поэтому целлюлоза (состоит из остатков глюкозы, связанныхβ- гликозидной связью) не переваривается.
Целлюлоза (клетчатка) относится к полисахаридам. Наряду с крахмалом
она является главным углеводом растений. Важнейшим полисахаридом человека, также построенным из остатков глюкозы, является гликоген. Крахмал и гликоген представлены разветвлёнными цепями глюкозы.
По химическому строению целлюлоза,
крахмал и гликоген являются гомополисахаридами (структура гликогена описана ниже)
Гетерополисахариды
представлены мукополисахаридами, протеогликанами и гликопротеинами .
studfiles.net